www.fimpolgy.com/JTF
The Journal of Theoretical Fimpology. Volume 1, Issue 2: e-20120609-1-2-3. May 18, 2013 (www.fimpology.com)
Six Models of Pregnancy-Associated Eukaryotic Cell Transmission among Fetus, Mother and Infant
Shu-dong Yin
ORCID ID: https://orcid.org/0009-0005-8661-6889
Cory H. E. R. & C. Inc., Burnaby, British Columbia, Canada
Pregnancy-associated cell transmission among fetus, mother and infant has attracted more and more attention during the past decades, among which, fetus-to-maternal cell transmission (FMCT) and maternal-to-fetal cell transmission (MFCT) are the two most studied. Recently, a novel model called fetus-breastmilk-breastfeeding-infant-cells cycle (FBBIC Cycle) has been proposed, which uncovered an unrecognized embryonic/fetal cell migration for the transmission of eukaryotic cells from fetus to infant him/herself via breastmilk and breastfeeding. Up to now, total six models: (i) FMCT, (ii) MFCT, (iii) gestation-associated fetus-to-fetus cell transmission (FFCT), (iv) breastmilk and breastfeeding-mediated maternal-to-infant cell transmission (MICT), (v) breastmilk and breastfeeding-mediated fetus-to-infant allogenic or inter-sibling cell transmission (FICT), and (vi) FBBIC Cycle constitute a complex system of eukaryotic cell exchange and interaction among mother, fetus and suckling infant during gestation and lactation. Among these pregnancy-associated models, FBBIC Cycle should be the major melody because only it reflects the most benefit of offspring during his/her prenatal and postnatal life in the physiological and immunological perspective without potential disadvantages introduced by allogenic and semi-allogenic cells. A thoroughly theoretical exploration in fimpology is essential to accurately interpreting prenatal diagnosis, making further development of molecular diagnostic techniques and deeply elucidating tolerogenic mechanisms in transplantation immunology.
Key Words: cell transmission; pregnancy; breastfeeding; FBBIC Cycle; microchimerism; maternal cells; fetal cells; stem cells; milkcells
Based on the existing data from studies on humans and non-human mammals, six models of eukaryotic cell transmission among mother, fetus and infant during gestation and lactation have been proposed although some need further direct supporting evidences: (i) gestation-associated embryonic/fetal cell migration into maternal body (FMCT), which results in fetal-maternal microchimerism (FMMC) (64,79,80,139-141); (ii) gestation-associated maternal cell migration into fetal body (MFCT), which results in maternal-to-fetal microchimerism (MFMC) (28,34,36,85,142,143); (iii) gestation-associated allogenic or inter-sibling fetal cell transmission between different fetuses (FFCT), or called "transmaternal sibling microchimerism" which results in fetofetal microchimerism (FFMC) (94-100,104); (iv) gestation and lactation-associated maternal host cell migration into the body of suckling infant via breastmilk and breastfeeding (MICT), which results in maternal-infantile microchimerism (MIMC) (30,84,89,106,110,111,144,145); (v) gestation and lactation-associated allogenic or inter-sibling fetal cell transmission to the suckling infant via breastmilk and breastfeeding (FICT), which results in fetoinfantile microchimerism (FIMC); and (vi) gestation and lactation associated fetus-to-infant his/her own fetal cell external transmission via breastmilk and breastfeeding, or called Fetus-Breastmilk-Breastfeeding-Infant-Cells Cycle (FBBIC Cycle) (82) (see Figure).
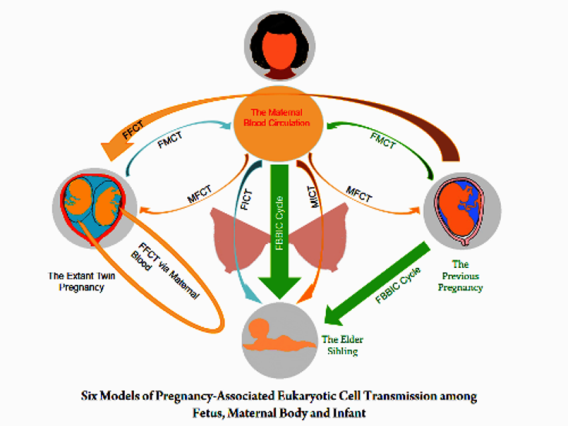
Figure: In the imaged mother and her offspring, gestation-associated embryonic/fetal cell migration into maternal body (FMCT) and gestation-associated maternal cell migration into fetal body (MFCT) may take place between the prenatal period of the first born offspring and his/her maternal body, and between the extant fetal twins and their maternal body. Maternal host cell migration into the body of suckling infant (MICT), allogenic or inter-sibling fetal cell transmission to the suckling infant (FICT), and fetus-to-infant his/her own fetal cell external transmission (FBBIC Cycle) may arise between the lactating mother and her suckling infants via breastfeeding. In addition, gestation-associated allogenic or inter-sibling fetal cell migration (FFCT) may take place when residual embryonic/fetal cells of the previous pregnancy migrate into the body of the extant fetal twins via the maternal blood circulation; and moreover, FFCT also exists between twin fetuses.
Interestingly, FMCT, MFCT, FFCT, MICT and FICT share a common characteristic that is the migration of foreign cells into tissues and/or organs of a macroorganism body. MFCT, FMCT, and FFCT occur during gestation, and MICT and FICT may take place postnatally in those infants who were breastfed with their own mothers' breastmilk, or with breastmilk from allogenic lactating mothers or donors.
In mammals including human beings, there are three major naturally connected physiological proceedings for producing offspring — (i) embryonic/fetal growth and development, (ii) production of breastmilk, and (iii) breastfeeding offspring. However, MFCT, FMCT, FFCT, MICT and FICT fail to reflect the physiological importance and the biological necessity for the continuity of these natural events. Only can FBBIC Cycle satisfy the most benefit of offspring during their prenatal and postnatal life in the physiological and immunological perspective, and therefore, should be recognized as the major physiological mechanism in these models.
In this manuscript, these different models of pregnancy-associated prenatal and postnatal cell transmission are briefly summarized.
I. Migration of Embryonic/Fetal Cells into Maternal Body (FMCT)
Migration of fetal cells into maternal organs and tissues (FMCT), or called fetomaternal microchimerism (FMMC) was the earliest recognized consequence of cellular migration from fetus to mother, and was first described by Georg Schmorl in a pathological case in 1893 (58,59). Studies on humans and non-human mammals indicated that migration of fetal cells into maternal blood is a normal physiological phenomenon that occurs in all pregnancies (15,59-61).
Detection of fetal cells in maternal body
The earliest presence of fetal cell in maternal body varied in different species. In murine model, fetal microchimerism begins soon after implantation, and increases with gestational age (9,15,16,18,23,60,62). In humans, the earliest detectable fetal cells in the body of normal pregnant women reported by different research groups was at 4-5 week- (3), 6 week- (63), and 15 week-gestational age (64) respectively.
Migration of fetal cell into maternal organs/tissues in humans appeared to increase with gestational age (24). The number of fetal cells in maternal blood varied widely ranging from one fetal cell per 10[4] to one fetal cell per 10[9] maternal cells (6). Lo and colleagues even showed that at least 17% of maternal cellular component was positive for fetal DNA (65).
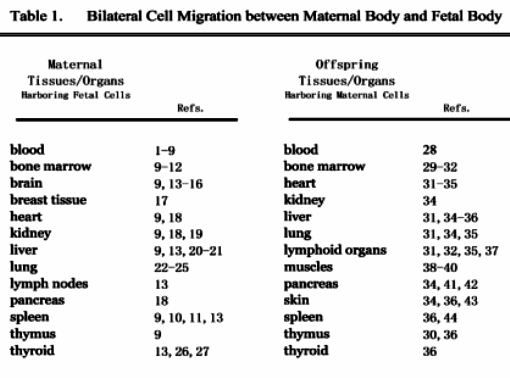
Many maternal tissues and organs of humans and some non-human mammal models were showed to harbor fetal cells (see Table 1); and moreover, those co-existing fetal cells were found to be multiple cell types including endothelial cells, mesenchymal stem cells /progenitors and neuronal cells in the relevant maternal tissues and organs (see Table 2).
Clinical significance and potential challenge
The potential physiological significance and pathological consequence of FMCT has been one of reasons for attracting researchers' attention during the past two decades (55,59,63,66-68). The existing literatures indicted that fetal cells seemed to involve in the pathogenesis of some clinical diseases, such as preeclampsia, rheumatoid arthritis, systemic lupus erythematosus (SLE) and systemic sclerosis during pregnancy (7,13,66,67,69-74), or possibly to play a role in maternal tissue repair during and after gestation (13,22,26,63,75,76). O'Donoghue even proposed that the co-existence of fetal cells in maternal body may be one of answers for the question of "Why is maternal life longer than paternal life?" (78).
The possible role played by fetal cells in anti-breast cancer attracted Gadi and colleagues' attention (79-81). However, while elucidating the anti-breast-cancer hypothesis, Gadi revealed that fetal cells co-existed in normal breast tissue of parous healthy women (17), which, to the best of my knowledge, may be the first direct supporting evidence of the FBBIC Cycle theory (82).
Another reason for study on FMCT is its potential value in prenatal genetic diagnosis (4,8,78). However, upon possible FFCT and FMCT from silent aborted pregnancies or elder siblings, how to interpret properly the results of prenatal genetic diagnosis is a challenge although Sato and colleagues showed that after abortion, a quick decline of fetal cells in the maternal blood circulation to an undetectable level needed only one month (83).
II. Migration of Maternal Cells into Fetal Body (MFCT)
Gestational maternal immunity transmission to fetus was described as early as in 1892 by Paul Erlich using experimental mouse models (59,84). Although the distinction between humoral and cellular immunities hadn't established until the emergence of modern immunology during the second half of the 20th century, some researchers believed that in Paul Erlich's mouse models, the transmission of maternal cell-mediated cellular immunity to fetus should be included (59,84). Maloney and colleagues further showed that maternal cells even persisted in the body of healthy offspring for as long as 49 years (59,85), which, therefore, has been recognized as a physiological phenomenon (59). Studies on humans and some non-human mammals showed that maternal cells were found in multiple tissues and organs of fetal, neonatal and adult offspring (see Table 1). Many maternal cell lineages that were found in offspring tissues and organs include erythrocytes, leukocytes (granulocytes, T and B lymphocytes) and platelets (see Table 2).
Clinical significance
The first impression of maternal cells in the body of offspring was associated with their pathological role (59,86,87). Microchimeric maternal cells in offspring may involve in the pathogenesis of some pediatric diseases, such as neonatal lupus syndrome (34,68,73), juvenile dermatomyositis (68,88), and type 1 diabetes (41). Moreover, the migration of maternal cells into fetal body was considered to be a prenatal mechanism for the vertical transfer of viral pathogens (89,90).
Maternal-donor related renal transplant
As early as in 1950s, animal experiments and clinical studies showed that maternal eukaryotic cells within fetal body may involve in tolerance genesis (91). The tolerance to noninherited maternal antigens (NIMAs) through the exposure to maternal cells in fetal body was proposed as one of tolerogenic mechanisms (89,92,93). However, in mammals including human beings, the exposure to NIMAs may occur during gestation and/or lactation via the following routes: (i) molecular exchange between mother and fetus, and/or between lactating mother and her suckling infant, and (ii) migration of maternal cells into fetal body, and/or migration of maternal cells into the body of her suckling infant, which depends on whether the infant being breastfed by his/her own mother. Therefore, at the cellular level, the tolerance to NIMAs may occur as early as during gestation via MFCT. Postnatal MICT may play a role in strengthening the tolerance to NIMAs. More discussion is in the following MICT section.
III. Gestation-Associated Fetal Cell Transmission between Different Fetuses (FFCT)
In the gestational fetus-to-fetus fetal cell transmission (FFCT) model, "Fetuses" are different individuals, and fetal cells from one fetus can co-exist within fetal body/bodies of the other/others. FFCT could occur theoretically in the following pregnant circumstances: (i) multiple pregnancies including twin pregnancy (94-98), in which, more than one fetus are usually growing and developing simultaneously within one uterus; (ii) residual fetal cells in maternal body from previously aborted embryo enter the body of another fetus of next pregnancy (99,100); and (iii) remaining fetal cells in maternal body from previously born brothers and/or sisters migrate into fetal body of younger brothers and/or sisters (101,102).
It is worth to point out that FFCT in the existing literatures of animal study on microchimerism has been often overlooked. In fact, many non-human mammal models used in experimental studies are multiple pregnancy animals, such as mice, rats, and pigs (103,104); and therefore, theoretically, there should be a fetus-to-fetus cell transmission or exchange during their normal gestation, which was supported by the latest laboratory evidences from studies on porcine littermates (103,104).
In humans, despite normal fetus-to-fetus microchimerism in plural pregnancy (94,98), there may be a FFCT in the extant pregnancy of multiparous women who previously experienced a silent or identified pregnancy. Nelson's group revealed an interesting phenomenon that male DNA or cells were often detected in healthy women who had never given birth to a son (99,100), for which, FFCT was discussed to be one of possible explanations by the authors (99).
Clinical significance
The inherited paternal antigens (IPAs) of one fetus in previous/or extant pregnancy may be noninherited paternal antigens (NIPA) to the other fetus of next/or extant pregnancy. The clinical significance of FFCT is elucidated in a noninherited-paternal-antigens-centric model for understanding transplantation immunology (see the article entitled "A Noninherited-Paternal-Antigens-Centric Model for the Birth Order and Sibship Size Effect in Transplantation Immunology" in this issue).
Offspring may contact NIPA via prenatal FFCT and postnatal FICT. Roelen and colleagues showed that no differences in the frequencies of cytotoxic T cell precursors (CTLp) and IL-2-producing helper T cell precursors (HTLp) against the noninherited maternal and the paternal HLA-A and -B antigens, and moreover, similar frequencies of CTLp and HTLp were observed between breastfed and non-breastfed children (105), which suggested the efficiency of gestational exposure to NIPA via prenatal FFCT may be superior to that of lactational exposure to NIPA via breastmilk and breastfeeding-mediated FICT. Perhaps, the significance of exposure to NIPA via postnatal FICT is in strengthening NIPA-tolerogenesis.
IV. Migration of Maternal Cells into Infantile Body (MICT)
Migration of maternal host cells into suckling body via breastmilk and breastfeeding, which results in maternal-infantile microchimerism (MIMC), was supported by many laboratory evidences from experimental studies on non-human mammals, such as murine (30,84,106-109), lambs (107), and piglets (110,111).
Traditionally, it was accepted generally during the 20th century that milkcells consisted of multiple end-differentiated maternal cell lineages, such as mononuclear phagocytes, polymorphonuclear leukocytes, lymphocytes, colostral corpuscles, and ductal/luminal epithelial cells. Such notion was reflected in almost all of the studies around milk cells (30,107,109,112-116). Recently, various stem/progenitor cells in breastmilk of human beings and non-human mammals were also assumed to be from maternal mammary stem cells (117-119), which was supported by that similar stem/progenitor cell properties were described on human breast epithelial cell line (120-123). Therefore, as the consequence, on the one hand, none of previous studies on breastfeeding-mediated cell transmission to offspring had further checked milkcells' origins; on the other hand, relevant laboratory techniques for distinguishing milkcells from different individuals haven't been developed yet. However, in the recently proposed FBBIC Cycle theory, milkcells were hypothesized to be heterogeneous and derived from both maternal and fetal sources (82). According to the new understanding, the co-existence of partial milkcells within the body of a suckling infant may be the consequence of MICT, FICT and FBBIC Cycle.
Clinical significance
In 1980s, some researchers found that breastfed patients who were the recipients of maternal-donor related renal transplant had better graft function rates than did non-breastfed counterparts (124,125), in which, the tolerance to noninherited maternal antigens (NIMAs) through the exposure to maternal cells via prenatal MFCT and postnatal MICT via breastfeeding was believed to be one of tolerogenic mechanisms (89,92,126).
However, comparing to MFCT, the real significance of MICT in the tolerance genesis to NIMAs needs further evaluation. Breastfeeding-mediated postnatal MICT may play a role as strengthening the tolerance to NIMAs as the strengthening immunization to a vaccine.
V. Migration of Sibling and Allogeneic Fetal Cells into Infantile Body (FICT)
Inter-sibling or allogeneic fetal cells transmission is defined to be a small amount of foreign fetal cells migrated into tissues or circulation of infants during lactation via breastfeeding, in which, "fetus" that fetal cells derived from and "infant" who harbored the fetal cells are not the same individual; and moreover, the fetal cells are not from the maternal host source.
It is not difficult to understand that the following three conditions were most likely to allow FICT to occur in a suckling infant: (i) the infant was breastfed by an allogeneic lactating mother; (ii) the infant was fed with donor breastmilk from Human Milk Bank; and (iii) the infant was breastfed by his/her own multiparous mother. In the above first two conditions, the obtained fetal cells by breastfed infant are allogeneic. In the third condition, the fetal cells left in breastmilk were derived from his or her siblings in previous pregnancies/abortions.
Theoretically, FICT and FBBIC Cycle may even occur simultaneously in infants who were breastfed by their own multiparous mothers. In other words, when breastfed by his/her own multiparous mother, suckling baby may regain two kinds of fetal cells: one is his or her own fetal cells defined under the FBBIC Cycle, and the other fetal cells left by his or her siblings in previous pregnancies/abortions. Even the possibility that FICT of some embryonic/fetal stem cells derived from maternal siblings and grandmother cannot be ruled out.
Embryonic stem cells were referred to those cells that come from an organism at its earliest stages of development (127), and can infinitely renew themselves and transform into multicellular lineages in the body of macroorganisms, such as human beings and non-human mammals (22,127).
Therefore, embryonic/fetal cells including stem cells in breastmilk theoretically may come from the following possible origins: (i) embryo/fetus of the extant pregnancy, (ii) embryonic/fetal siblings in extant multiple pregnancy, (iii) embryo/fetus of previously aborted or successful pregnancies, (iv) embryo/fetus of maternal siblings, (v) mother herself, and (vi) grandmother.
Clinical significance
To the best of my knowledge, it is the first time to theoretically propose that there may a FICT mechanism that may mediate sibling embryonic/fetal cells and allogeneic embryonic/fetal cells in breastmilk into the body of suckling infants via breastfeeding.
FICT may involve in tolerogenic mechanisms of transplantation immunology, which has been discussed in the article entitled "A noninherited-paternal-antigens-centric model for the birth order and sibship size effect in transplantation immunology" in this issue.
VI. Fetus-to-Infant His/Her Own Cells Transmission Cycle or Fetus-Breastmilk-Breastfeeding-Infant-Cells Cycle (FBBIC Cycle)
Recently, a model called Fetus-Breastmilk-Breastfeeding-Infant-Cells Cycle (FBBIC Cycle) was proposed for revealing an unrecognized mechanism (82), in which, "fetus" and "infant" in this model are the same individual, and moreover, the absorbed fetal cells from breastmilk were actually derived from the suckling infant's own host cells. Therefore, FBBIC Cycle reflects the fetus-to- infant his/her own cell external transmission via breastmilk and breastfeeding.
If an infant has never been breastfed by his/her own mother's breastmilk, his/her FBBIC Cycle may be missed (82). In fact, among the six different eukaryotic cell transmissions, only is FBBIC Cycle able to account for why the following three physiological events should be naturally connected: (i) embryonic/fetal growth and development, (ii) production of breastmilk and (iii) breastfeeding offspring.
Pregnancy-associated maternal immune protection blank (MIPB)
In humans, the gestational course from a fertilized egg to a matured fetus normally needs 37-40 weeks. In clinical medicine, pregnancies of less than 37 weeks or longer than 42 weeks are usually defined as a pathological status. During gestation, maternal body continuously contacts novel external foreign antigens including those in foods and air. It is well known that the establishment of immune to a specific antigenic stimulation usually needs 4 weeks.
During gestation, newly formed maternal immunity cannot be transferred to her offspring of extant pregnancy via DNA inheritance. However, such newly acquired maternal immune protection can be transferred to her offspring mainly via trans-placental IgG and possibly gestational MFCT. Therefore, the gestational period is a relative maternal immune protection blank (MIPB) for novel external foreign antigens for offspring, in which, there is a 4-week absolute MIPB for gestational offspring.
How does offspring overcome this MIPB period? FBBIC Cycle may be one of mechanisms for answering this question. During gestation, fetal cells migrate into the maternal blood circulation and other tissues/organs where they have an opportunity to contact the novel foreign antigens that the pregnant maternal body is experiencing. Normal pregnancy has been revealed to be a systemic Th2 characterized immune response in maternal body since 1993 (128). Moreover, studies on human breastmilk have already showed that there is certain amount of microorganisms in normal breastmilk including bacterial (129-132), fungal (146), and viral species (133-138), and therefore, any episodes of exposure to novel bacterial, viral and fungal agents or other foreign antigens during gestation may affect both maternal immune cells and those microchimeric fetal cells. Because some fetal cells are embryonic stem cells, and have the capability to differentiate into various cell lineages including immune cells (such as T memory cells, for example). If these fetal cells return the body of suckling infant via breastfeeding, doubtlessly, the acquired cellular immunity during gestation in maternal body will be transferred to postnatal suckling neonates.
It already has been proposed that fetal cell-derived maternal alloantigens-tolerogenic fetal T regulatory cells (MATF-TRCs) (35,37) and paternal alloantigens-tolerogenic fetal T regulatory cells (PATF-TRCs) (82) may involve in suppressing postnatal antimaternal or antipaternal immune reaction to maternal or paternal graft (82).
Conclusion
Eukaryotic cell transmission among fetus, mother and infant during pregnancy is a complex process, which covers both gestation and lactation. Up to now, total six pregnancy-associated cell transmission models have been proposed, among which, FBBIC Cycle is the only model that can satisfy the most benefit of offspring's prenatal and postnatal life in physiology and immunology, and account for the necessity and the continuity of gestation to lactation in pregnancy (82). Prenatal FFCT and postnatal FICT may involve in paternal alloantigens-tolerogenic mechanism of offspring. A thoroughly theoretical exploration in the quadruple relationship of fetus, infant, mother and father is one of the current major concerns of fimpology, which doubtlessly will contribute to interpretation of prenatal genetic diagnosis, development of molecular diagnostic techniques and elucidation of tolerogenic mechanisms in transplantation immunology.
Abbreviations
FMCT: gestation-associated fetus-to-maternal cell transmission;
MFCT: gestation-associated maternal-to-fetal cell transmission;
FFCT: gestation-associated fetus-to-fetus cell transmission;
MICT: breastmilk and breastfeeding-mediated maternal-to-infant cell transmission;
FICT: breastmilk and breastfeeding-mediated transmission of sibling and allogenic fetal cells into infantile body;
FBBIC Cycle: fetus-to-infant his/her own fetal cell external transmission via breastfeeding;
FMMC: fetal-maternal microchimerism;
MFMC: maternal-to-fetal microchimerism;
FFMC: fetofetal microchimerism;
MIMC: maternal-infantile microchimerism;
FIMC: fetoinfantile microchimerism;
NIPAs: noninherited paternal antigens;
NIMAs: noninherited maternal antigens.
Reference
1. Schroder J, De la Chapelle A. Fetal lymphocytes in the maternal blood. Blood 1972; 39(2): 153-62
2. Schroder J, Tiilikainen A, De la Chapelle A. Fetal leukocytes in the maternal circulation after delivery. I. Cytological aspects. Transplantation 1974; 17(4): 346-54
3. Thomas MR, Williamson R, Craft I, Yazdani N, Rodeck CH. Y chromosome sequence DNA amplified from peripheral blood of women in early pregnancy. Lancet 1994; 343(8894): 413-4
4. Ariga H, Ohto H, Busch MP, Imamura S, Watson R, Reed W, et al. Kinetics of fetal cellular and cell-free DNA in the maternal circulation during and after pregnancy: implications for noninvasive prenatal diagnosis. Transfusion 2001; 41(12): 1524-30
5. Toda I, Kuwana M, Tsubota K, Kawakami Y. Lack of evidence for an increased microchimerism in the circulation of patients with Sjögren's syndrome. Ann Rheum Dis 2001; 60(3): 248-53
6. Krabchi K, Gros-Louis F, Yan J, BronsardM, Masse J, Forest JC, et al. Quantification of all fetal nucleated cells in maternal blood between the 18th and 22nd weeks of pregnancy using molecular cytogenetic techniques. Clin Genet. 2001; 60(2): 145-50
7. Lambert NC, Lo YM, Erickson TD, Tylee TS, Guthrie KA, Furst DE, et al. Male microchimerism in healthy women and women with scleroderma: cells or circulating DNA? a quantitative answer. Blood 2002; 100(8): 2845-51
8. O'Donoghue K, Choolani M, Chan J, de la Fuente J, Kumar S, Campagnoli C, et al. Identification of fetal mesenchymal stem cells in maternal blood: implications for non-invasive prenatal diagnosis. Mol Hum Reprod 2003; 9(8): 497-502
9. Fujiki Y, Johnson KL, Peter I, Tighiouart H, Bianchi DW. Fetal cells in the pregnant mouse are diverse and express a variety of progenitor and differentiated cell markers. Biol Reprod 2009; 81(1): 26-32
10. Liegeois A, Gaillard MC, Ourve E, Lewin D. Microchimerism in pregnant mice. Transplant Proc 1981; 13(1 Pt 2): 1250-2
11. Philip PJ, Ayraud N, Masseyeff R. Transfer, tissue localization and proliferation of fetal cells in pregnant mice. Immunol Lett 1982; 4(3): 175-8
12. O'Donoghue K, Chan J, de la Fuente J, Kennea N, Sandison A, Anderson JR, et al. Microchimerism in female bone marrow and bone decades after fetal mesenchymal stem-cell trafficking in pregnancy. Lancet 2004; 364(9429):179-82
13. Khosrotehrani K, Johnson KL, Cha DH, Salomon RN, Bianchi DW. Transfer of fetal cells with multilineage potential to maternal tissue. JAMA 2004; 292(1): 75-80
14. Tan XW, Liao H, Sun L, Okabe M, Xiao ZC, Dawe GS. Fetal microchimerism in the maternal mouse brain: a novel population of fetal progenitor or stem cells able to cross the blood-brain barrier? Stem Cells 2005; 23(10):1443–52
15. Dawe GS, Tan XW, Xiao ZC. Cell migration from baby to mother. Cell Adh Migr 2007; 1(1): 19-27
16. Sunami R, Komuro M, Yuminamochi T, Hoshi K, Hirata S. Fetal cell microchimerism develops through the migration of fetus-derived cells to the maternal organs early after implantation. J Reprod Immunol 2010; 84(2): 117-23
17. Gadi VK. Fetal microchimerism in breast from women with and without breast cancer. Breast Cancer Res Treat. 2010; 121(1): 241-4. doi: 10.1007/s10549-009-0548-1) (PMID: 19768535
18. Sunami R, Komuro M, Tagaya H, Hirata S. Migration of microchimeric fetal cells into maternal circulation before placenta formation. Chimerism 2010; 1(2): 66-8
19. Wang Y, Iwatani H, Ito T, Horimoto N, Yamato M, Matsui I, et al. Fetal cells in mother rats contribute to the remodeling of liver and kidney after injury. Biochem Biophys Res Commun 2004; 325(3): 961-7
20. Tanaka A, Lindor K, Gish R, Batts K, Shiratori Y, Omata M, et al. Fetal microchimerism alone does not contribute to the induction of primary biliary cirrhosis. Hepatology 1999; 30(4): 833-8
21. Invernizzi P, De Andreis C, Sirchia SM, Battezzati PM, Zuin M, Rossella F, et al. Blood fetal microchimerism in primary biliary cirrhosis. Clin Exp Immunol 2000; 122(3): 418-22
22. O'Donoghue K, Sultan HA, Al-Allaf FA, Anderson JR, Wyatt-Ashmead J, Fisk NM. Microchimeric fetal cells cluster at sites of tissue injury in lung decades after pregnancy. Reprod Biomed Online 2008; 16(3): 382-90
23. Fujiki Y, Johnson KL, Tighiouart H, Peter I, Bianchi DW. Fetomaternal trafficking in the mouse increases as delivery approaches and is highest in the maternal lung. Biol Reprod 2008; 79(5): 841-8
24. Benirschke K, Willes L. Deportation of trophoblastic emboli to maternal lung: A source of cell-free DNA in maternal blood? Chimerism 2010; 1(1): 15-8
25. Johnson KL, Tao K, Stroh H, Kallenbach L, Peter I, Richey L, et al. Increased fetal cell trafficking in murine lung following complete pregnancy loss from exposure to lipopolysaccharide. Fertil Steril 2010; 93(5): 1718-21
26. Srivatsa B, Srivatsa S, Johnson KL, Samura O, Lee SL, Bianchi DW. Microchimerism of presumed fetal origin in thyroid specimens from women: a case control study. Lancet 2001; 358(9298): 2034-8
27. Renn C, Ramos Lopez E, Steimle-Grauer SA, Ziolkowski P, Pani MA, Luther C, et al. Thyroid fetal male microchimerisms in mothers with thyroid disorders: presence of Y-chromosomal immunofluorescence in thyroid-infiltrating lymphocytes is more prevalent in Hashimoto's thyroiditis and Graves' disease than in follicular adenomas. J Clin Endocrinol Metab 2004; 89(11): 5810-4
28. Hall JM, Lingenfelter P, Adams SL, Lasser D, Hansen JA, Bean MA. Detection of maternal cells in human umbilical cord blood using fluorescence in situ hybridization. Blood 1995; 86(7): 2829-32
29. Marleau AM, Greenwood JD, Wei Q, Singh B, Croy BA. Chimerism of murine fetal bone marrow by maternal cells occurs in late gestation and persists into adulthood. Lab Invest 2003; 83(5): 673-81
30. Zhou L, Yoshimura Y, Huang Y, Suzuki R, Yokoyama M, Okabe M, et al. Two independent pathways of maternal cell transmission to offspring: through placenta during pregnancy and by breast-feeding after birth. Immunology 2000; 101(4): 570-80
31. Piotrowski P, Croy BA. Maternal cells are widely distributed in murine fetuses in utero. Biol Reprod 1996; 54(5): 1103-10
32. Dutta P, Burlingham WJ. Stem cell microchimerism and tolerance to non-inherited maternal antigens. Chimerism 2010; 1(1): 2-10
33. Stevens AM, Hermes HM, Rutledge JC, Buyon JP, Nelson JL. Myocardial-tissue-specific phenotype of maternal microchimerism in neonatal lupus congenital heart block. Lancet 2003; 362(9396): 1617-23
34. Stevens AM, Hermes HM, Kiefer MM, Rutledge JC, Nelson JL. Chimeric maternal cells with tissue-specific antigen expression and morphology are common in infant tissues. Pediatr Dev Pathol 2009; 12(5): 337-46
35. Dutta P, Molitor-Dart M, Bobadilla JL, Roenneburg DA, Yan Z, Torrealba JR, et al. Microchimerism is strongly correlated with tolerance to noninherited maternal antigens in mice. Blood 2009; 114(17): 3578-87
36. Srivatsa B, Srivatsa S, Johnson KL, Bianchi DW. Maternal cell microchimerism in newborn tissues. J Pediatr 2003; 142(1): 31-5
37. Mold JE, Michaelsson J, Burt TD, Muench MO, Beckerman KP, Busch MP, et al. Maternal alloantigens promote the development of tolerogenic fetal regulatory T cells in utero. Science 2008; 322(5907): 1562-5
38. Artlett CM, Ramos R, Jiminez SA, Patterson K, Miller FW, Rider LG, et al. Chimeric cells of maternal origin in juvenile idiopathic inflammatory myopathies. Lancet 2000; 356(9248): 2155-6
39. Reed AM, Picornell YJ, Harwood A, Kredich DW. Chimerism in children with juvenile dermatomyositis. Lancet 2000; 356(9248): 2156-7
40. Ye Y, van Zyl B, Varsani H, Wedderburn LR, Ramanan A, Gillespie KM; Juvenile Dermatomyositis Research Group. Maternal microchimerism in muscle biopsies from children with juvenile dermatomyositis. Rheumatology (Oxford) 2012; 51(6): 987-91
41. Nelson JL, Gillespie KM, Lambert NC, Stevens AM, Loubiere LS, Rutledge JC, et al. Maternal microchimerism in peripheral blood in type 1 diabetes and pancreatic islet beta cell microchimerism. Proc Natl Acad Sci USA 2007; 104(5): 1637-42
42. Vanzyl B, Planas R, Ye Y, Foulis A, de Krijger RR, Vives-Pi M, et al. Why are levels of maternal microchimerism higher in type 1 diabetes pancreas? Chimerism 2010; 1(2): 45-50
43. McNallan KT, Aponte C, el-Azhary R, Mason T, Nelson AM, Paat JJ, et al. Immunophenotyping of chimeric cells in localized scleroderma. Rheumatology (Oxford) 2007; 46[3): 398-402
44. Shimamura M, Ohta S, Suzuki R, Yamazaki K. Transmission of maternal blood cells to the fetus during pregnancy: Detection in mouse neonatal spleen by immunofluorescence flow cytometry and polymerase chain reaction. Blood 1994; 83(4): 926-30
45. Nguyen Huu S, Oster M, Uzan S, Chareyre F, Aractingi S, Khosrotehrani K. Maternal neoangiogenesis during pregnancy partly derives from fetal endothelial progenitor cells. Proc Natl Acad Sci USA 2007; 104(6): 1871-6
46. Parant O, Dubernard G, Challier JC, Oster M, Uzan S, Aractingi S, et al. CD34+ cells in maternal placental blood are mainly fetal in origin and express endothelial markers. Lab Invest 2009; 89(8): 915-23
47. Lo YMD, Morey AL, Wainscoat JS, Fleming KA. Culture of fetal erythroid cells from maternal peripheral blood. Lancet 1994; 344(8917): 264-5
48. Lewis DE, Schober W, Murrell S, Nguyen D, Scott J, Boinoff J. Rare event selection of fetal nucleated erythrocytes in maternal blood by flowcytometry. Cytometry 1996; 23(3): 218-27
49. Valerio D, Aiello R, Altieri V, Malato AP, Fortunato A, Canazio A, et al. Culture of fetal erythroid progenitor cells from maternal blood for non-invasive prenatal genetic diagnosis. Prenat Diagn 1996; 16(12): 1073-82
50. Brazelton T, Rossi F, Keshet G, Blau HM. From marrow to brain: expression of neuronal phenotypes in adult mice. Science 2000; 290(5497): 1775-9
51. Krause DS, Theise ND, Collector MI, Henegariu O, Hwang S, Gardner R, et al. Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell 2001; 105(3): 369-77
52. Walknowska J, Conte FA, Grumbach MM. Practical and theoretical implications of fetal-maternal lymphocyte transfer. Lancet 1969; 1(7606): 1119-22
53. Schindler AM, Martin-du-Pan R. Prenatal diagnosis of fetal lymphocytes in the maternal blood. Obstet Gynecol 1972; 40(3): 340-6
54. Assche EL, Susanne C. Increase in the amount of fetal lymphocytes in maternal blood during pregnancy. J Med Genet 1980; 17(4): 267-72
55. Sunku Cuddapah CS, Gadi VK, de Laval de Lacoste B, Guthrie KA, Nelson JL. Maternal and fetal microchimerism in granulocytes. Chimerism 2010; 1: 11-4
56. Sargent IL, Johansen M, Chau S, Redman CWG. Clinical experience: Isolating trophoblasts from maternal blood. Ann N Y Acad Sci 1994; 731: 154-61
57. van Wijk IJ, van Vugt JM, Mulders MA, Konst AA, Weima SM, Oudejans CB. Enrichment of fetal trophoblast cells from the maternal peripheral blood followed by detection of fetal deoxyribonucleic acid with a nested X/Y polymerase chain reaction. Am J Obstet Gynecol 1996; 174(3): 871-8
58. Lapaire O, Holzgreve W, Oosterwijk JC, Brinkhaus R, Bianchi DW. Georg Schmorl on trophoblasts in the maternal circulation. Placenta 2007; 28(1): 1-5
59. Gammill HS, Nelson JL. Naturally acquired microchimerism. Int J Dev Biol 2010; 54(2-3): 531-43
60. Khosrotehrani K, Johnson KL, Guegan S, Stroh H, Bianchi DW. Natural history of fetal cell microchimerism during and following murine pregnancy. J Reprod Immunol 2005; 66(1): 1-12
61. Adams Waldorf KM, Gammill HS, Lucas J, Aydelotte TM, Leisenring WM, Lambert NC, et al. Dynamic changes in fetal microchimerism in maternal peripheral blood mononuclear cells, CD4+ and CD8+ cells in normal pregnancy. Placenta 2010; 31(7): 589-94
62. Lee ESM, Bou-Gharios G, Seppanen E, Khosrotehrani K, Fisk NM. Fetal stem cell microchimerism: natural-born healers or killers? Mol Hum Reprod 2010; 16(11): 869-78
63. Santos MA, O'Donoghue K, Wyatt-Ashmead J, Fisk NM. Fetal cells in the maternal appendix: a marker of inflammation or fetal tissue repair? Hum Reprod 2008; 23(10): 2319-25
64. Herzenberg LA, Bianchi DW, Schroder J, Cann HM, Iverson GM. Fetal cells in the blood of pregnant women: detection and enrichment by fluorescence-activated cell sorting. Proc Natl Acad Sci USA 1979; 76(3): 1453-5
65. Lo YM, Corbetta N, Chamberlain PF, Rai V, Sargent IL, et al. Presence of fetal DNA in maternal plasma and serum. Lancet 1997; 350(9076): 485-7
66. Bianchi DW, Zickwolf GK, Weil GJ, Sylvester S, DeMaria MA. Male fetal progenitor cells persist in maternal blood for as long as 27 years postpartum. Proc Natl Acad Sci USA 1996; 93(2): 705-8
67. Artlett CM, Smith JB, Jimenez SA. Identification of fetal DNA and cells in skin lesions from women with systemic sclerosis. N Engl J Med 1998; 338(17): 1186-91
68. Ye Y, Berendine VZ, Hellmich C, Gillespie KM. Microchimerism: covert genetics? Int J Mol Epidemiol Genet 2010; 1(4): 350-7
69. Ruiz-Irastorza G, Lima F, Alves J, Khamashta MA, Simpson J, Hughes GR, et al. Increased rate of lupus flare during pregnancy and the puerperium: a prospective study of 78 pregnancies. Br J Rheumatol 1996; 35(2): 133-8
70. Nelson JL, Ostensen M. Pregnancy and rheumatoid arthritis. Rheum Dis Clin North Am 1997; 23(1): 195-212
71. Holzgreve W, Ghezzi E, DiNaro E, Ganshirt D, Maymon E, Hahn S. Fetomaternal cell traffic is disturbed in preeclampsia. Obstet Gynecol 1998; 91(5 Pt 1): 669-72
72. Johnson KL, Nelson JL, Furst DE, McSweeney PA, Roberts DJ, Zhen DK, et al. Fetal cell microchimerism in tissue from multiple sites in women with systemic sclerosis. Arthritis Rheum 2001; 44(8): 1848-54
73. Adams Waldorf KM, Nelson JL. Autoimmune disease during pregnancy and the microchimerism legacy of pregnancy. Immunol Invest 2008; 37(5): 631-44
74. Evans PC, Lambert N, Maloney S, Furst DE, Moore JM, Nelson JL. Long-term fetal microchimerism in peripheral blood mononuclear cell subsets in healthy women and women with scleroderma. Blood 1999; 93(6): 2033-7
75. Mikhail MA, M'Hamdi H, Welsh J, Levicar N, Marley SB, Nicholls JP, et al. High frequency of fetal cells within a primitive stem cell population in maternal blood. Hum Reprod. 2008; 23(4): 928-33
76. Rossi G. Nature of stem cells involved in fetomaternal microchimerism. Lancet. 2004; 364(9449): 1936
77. Srivatsa B, Srivatsa S, Johnson KL, Samura O, Lee SL, Bianchi DW. Microchimerism of presumed fetal origin in thyroid specimens from women: a case-control study. Lancet. 2001; 358(9298): 2034-8
78. O'Donoghue K. Fetal microchimerism and maternal health during and after pregnancy. Obstet Med 2008; 1(2): 56-64
79. Gadi VK, Nelson JL. Fetal microchimerism in women with breast cancer. Cancer Res 2007; 67(19): 9035-8 (PMID: 17909006)
80. Gadi VK, Malone KE, Guthrie KA, Porter PL, Nelson JL. Case-control study of fetal microchimerism and breast cancer. PLoS One. 2008; 3(3): e1706. (PMID: 18320027)
81. Gadi VK. Fetal microchimerism and cancer. Cancer Lett 2009; 276(1): 8-13. doi: 10.1016/j.canlet.2008.07.025.
82. Yin S-d. Fetus-breastmilk-breastfeeding-infant-cells cycle: fetus-to-infant his/her own fetal cell external transmission via breastfeeding. The Journal of Theoretical Fimpology. 2013; 1(1): e-20120612-2. Available from:
http://www.fimpology.com83. Sato T, Fujimori K, Sato A, Ohto H. Microchimerism after induced or spontaneous abortion. Obstet Gynecol 2008; 112(3): 593-7
84. Arvola M, Gustafsson E, Svensson L, Jansson L, Holmdahl R, Heyman B, et al. Immunoglobulin-secreting cells of maternal origin can be detected in B cell-deficient mice. Biol Reprod 2000; 63(6): 1817-24
85. Maloney S, Smith A, Furst DE, Myerson D, Rupert K, Evans PC, et al. Microchimerism of maternal origin persists into adult life. J Clin Invest. 1999; 104(1): 41-7
86. Reynolds AG. Placental metastasis from malignant melanoma; report of a case. Obstet Gynecol 1955; 6(2): 205-9
87. Pollack MS, Kapoor N, Sorell M, Kim SJ, Christinasen FT, Silver DM, et al. DR-positive maternal engrafted T cells in a severe combined immunodeficiency patient without-graft-versus-host disease. Transplantation 1980; 30(5): 331-4
88. Reed AM, McNallan K, Wettstein P, Vehe R, Ober C. Does HLA-dependent chimerism underlie the pathogenesis of juvenile dermatomyositis? J Immunol 2004; 172(8): 5041-6
89. Maurel M-C, Kanellopoulos-Langevin C. Heredity—venturing beyond genetics. Biol Reprod 2008; 79(1): 2-8
90. Schwartz DH, Sharma UK, Perlman EJ, Blakemore K. Adherence of human immunodeficiency virus-infected lymphocytes to fetal placental cells: a model of maternal fetal transmission. Proc Natl Acad Sci USA 1995; 92(4): 978-82
91. Billingham RE, Brent L, Medawar PB. Actively acquired tolerance of foreign cells. Nature 1953; 172(4379): 603-6
92. van Rood JJ, Claas F. Both self and noninherited maternal HLA antigens influence the immune response. Immunol Today. 2000; 21(6): 269-73
93. Andrassy J, Kusaka S, Jankowska-Gan E, Torrealba JR, Haynes LD, Marthaler BR, et al. Tolerance to noninherited maternal MHC antigens in mice. J Immunol. 2003; 171(10): 5554-61
94. van Dijk BA, Boomsma DI, de Man AJ. Blood group chimerism in human multiple births is not rare. Am J Med Genet. 1996; 61(3): 264-8
95. Shalev SA, Shalev E, Pras E, Shneor Y, Gazit E, Yaron Y, et al. Evidence for blood chimerism in dizygotic spontaneous twin pregnancy discordant for Down syndrome. Prenat Diagn. 2006; 26(9): 782-4
96. Sperling L, Kiil C, Larsen LU, Brocks V, Wojdemann KR, Qvist I, et al. Detection of chromosomal abnormalities, congenital abnormalities and transfusion syndrome in twins. Ultrasound Obstet Gynecol. 2007; 29(5): 517-26
97. Ekelund CK, Skibsted L, Sogaard K, Main KM, Dziegiel MH, Schwartz M, et al. Dizygotic monochorionic twin pregnancy conceived following intracytoplasmic sperm injection treatment and complicated by twin-twin transfusion syndrome and blood chimerism. Ultrasound Obstet Gynecol. 2008; 32(6): 832-4
98. Jang JH, Jung H, Kim JH, Park WS, Kim SH. Blood chimerism in a dizygotic dichorionic pregnancy. Korean J Lab Med. 2010; 30(5): 521-4
99. Lambert NC, Pang JM, Yan Z, Erickson TD, Stevens AM, Furst DE, et al. Male microchimerism in women with systemic sclerosis and healthy women who have never given birth to a son. Ann Rheum Dis. 2005; 64(6): 845-8
100. Yan Z, Lambert NC, Guthrie KA, Porter AJ, Loubiere LS, Madeleine MM, et al. Male microchimerism in women without sons: quantitative assessment and correlation with pregnancy history. Am J Med. 2005; 118(8): 899-906
101. Bucher C, Stern M, Buser A, Heim D, Paulussen M, Halter J, et al. Role of primacy of birth in HLA-identical sibling transplantation. Blood. 2007; 110(1): 468-9
102. Gratwohl A, Doehler B, Stern M, Bucher C, Passweg J, Opelz G. Birth order and outcome after HLA-identical sibling donor transplantation. Blood. 2009; 114(27): 5569-70
103. McConico A, Butters K, Lien K, Knudsen B, Wu X, Platt JL, et al. In utero cell transfer between porcine littermates. Reprod Fertil Dev. 2011; 23(2): 297-302
104. Karniychuk UU, Van Breedam W, Van Roy N, Rogel-Gaillard C, Nauwynck HJ. Demonstration of microchimerism in pregnant sows and effects of congenital PRRSV infection. Vet Res. 2012; 43(1): 19
105. Roelen DL, van Bree FP, van Beelen E, van Rood JJ, Claas FH. No evidence of an influence of the noninherited maternal HLA antigens on the alloreactive T cell repertoire in healthy individuals. Transplantation. 1995; 59(12): 1728-33
106. Ma LJ, Walter B, Deguzman A, Muller HK, Walker AM. Trans-epithelial immune cell transfer during suckling modulates delayed-type hypersensitivity in recipients as a function of gender. PLoS One. 2008; 3(10): e3562
107. Sheldrake RF, Husband AJ. Intestinal uptake of intact maternal lymphocytes by neonatal rats and lambs. Res Vet Sci. 1985; 39(1): 10-5
108. Weiler IJ, Hickler W, Sprenger R. Demonstration that milk cells invade the suckling neonatal mouse. Am J Reprod Immunol. 1983; 4(2): 95-8
109. Kumar SN, Stewart GL, Steven WM, Seelig LL Jr. Role of T cell subsets in the maternal-to-neonatal transmission of immunity against Trichinella spiralis during lactation in rats. J Reprod Immunol. 1990; 17(1): 69-78
110. Tuboly S, Bernath S, Glavits R, Medveczky I. Intestinal absorption of colostral lymphoid cells in newborn piglets. Vet Immunol Immunopathol. 1988; 20(1): 75-85
111. Williams PP. Immunomodulating effects of intestinal absorbed maternal colostral leukocytes by neonatal pigs. Can J Vet Res. 1993; 57(1): 1-8
112. Dorosko SM, Connor RI. Primary human mammary epithelial cells endocytose HIV-1 and facilitate viral infection of CD4+ T lymphocytes. J Virol. 2010; 84(20): 10533-42
113. Thompson PA, Kadlubar FF, Vena SM, Hill HL, McClure GH, McDaniel LP, et al. Exfoliated ductal epithelial cells in human breast milk: a source of target tissue DNA for molecular epidemiologic studies of breast cancer. Cancer Epidemiol Biomarkers Prev. 1998; 7(1): 37-42
114. Smith CW, Goldman AS. The cells of human colostrum. I. In vitro studies of morphology and functions. Pediatr Res. 1968; 2(2): 103-9
115. Ho FC, Wong RL, Lawton JW. Human colostral and breast milk cells. A light and electron microscopic study. Acta Paediatr Scand. 1979; 68(3): 389-96
116. Crago SS, Prince SJ, Pretlow TG, McGhee JR, Mestecky J. Human colostral cells. I. Separation and characterization. Clin Exp Immunol. 1979; 38(3): 585-97
117. Cregan MD, Fan Y, Appelbee A, Brown ML, Klopcic B, Koppen J, et al. Identification of nestin-positive putative mammary stem cells in human breastmilk. Cell Tissue Res. 2007; 329(1): 129-36
118. Fan Y, Chong YS, Choolani MA, Cregan MD, Chan JK. Unravelling the mystery of stem/progenitor cells in human breast milk. PLoS One. 2010; 5(12): e14421
119. Thomas E, Zeps N, Cregan M, Hartmann P, Martin T. 14-3-3σ (sigma) regulates proliferation and differentiation of multipotent p63-positive cells isolated from human breastmilk. Cell Cycle. 2011; 10(2): 278-84
120. Gudjonsson T, Villadsen R, Nielsen HL, Ronnov-Jessen L, Bissell MJ, Petersen OW. Isolation, immortalization, and characterization of a human breast epithelial cell line with stem cell properties. Genes Dev. 2002; 16(6): 693-706
121. Clayton H, Titley I, Vivanco M. Growth and differentiation of progenitor/stem cells derived from the human mammary gland. Exp Cell Res. 2004; 297(2): 444-60
122. Clarke RB, Spence K, Anderson E, Howell A, Okano H, Potten CS. A putative human breast stem cell population is enriched for steroid receptor-positive cells. Dev Biol. 2005; 277(2): 443-56
123. Lu X, Li H, Xu K, Nesland JM, Suo Z. MUC-1-/ESA+ progenitor cells in normal, benign and malignant human breast epithelial cells. Histol Histopathol. 2009; 24(11): 1381-90
124. Kois WE, Campbell DA Jr, Lorber MI, Sweeton JC, Dafoe DC. Influence of breast feeding on subsequent reactivity to a related renal allograft. J Surg Res. 1984; 37(2): 89-93
125. Campbell DA Jr, Lorber MI, Sweeton JC, Turcotte JG, Niederhuber JE, Beer AE. Breast feeding and maternal-donor renal allografts. Possibly the original donor-specific transfusion. Transplantation. 1984; 37(4): 340-4
126. Andrassy J, Kusaka S, Jankowska-Gan E, Torrealba JR, Haynes LD, Marthaler BR, et al. Tolerance to noninherited maternal MHC antigens in mice. J Immunol. 2003; 171(10): 5554-61
127. Shevde N. Stem cells: flexible friends. Nature. 2012; 483(7387): S22-6
128. Wegmann TG, Lin H, Guilbert L, Mosmann TR. Bidirectional cytokine interactions in the maternal-fetal relationship: is successful pregnancy a TH2 phenomenon? Immunol Today. 1993; 14(7): 353-6
129. Heikkila MP, Saris PE. Inhibition of Staphylococcus aureus by the commensal bacteria of human milk. J Appl Microbiol. 2003; 95(3): 471-8
130. Martin R, Langa S, Reviriego C, et al. Human milk is a source of lactic acid bacteria for the infant gut. J Pediatr. 2003; 143(6): 754-8
131. Lindemann PC, Foshaugen I, Lindemann R. Characteristics of breast milk and serology of women donating breast milk to a milk bank. Arch Dis Child Fetal Neonatal Ed . 2004; 89(5): F440-1
132. Perez PF, Dore J, Leclerc M, Levenez F, Benyacoub J, Serrant P, et al. Bacterial imprinting of the neonatal immune system: lessons from maternal cells? Pediatrics. 2007; 119(3): e724-32
133. Junker AK, Thomas EE, Radcliffe A, Forsyth RB, Davidson AG, Rymo L. Epstein-Barr virus shedding in breast milk. Am J Med Sci. 1991; 302(4): 220-3
134. Kotronias D, Kapranos N. Detection of herpes simplex virus DNA in maternal breast milk by in situ hybridization with tyramide signal amplification. In Vivo. 1999; 13(6): 463-6
135. Meier J, Lienicke U, Tschirch E, Kruger DH, Wauer RR, Prosch S. Human cytomegalovirus reactivation during lactation and mother-to-child transmission in preterm infants. J Clin Microbiol. 2005; 43(3): 1318-24
136. Murata H, Nii R, Ito M, Ihara T, Komada Y. Quantitative detection of HCMV-DNA in saliva from infants and breast milk on real-time polymerase chain reaction. Pediatr Int. 2009; 51(4): 530-4
137. Lawson JS, Heads J, Glenn WK, Whitaker NJ. Breastfeeding, breast milk and viruses. BMC Womens Health. 2007; 7(): 17
138. Sarkola M, Rintala M, Grenman S, Syrjanen S. Human papillomavirus DNA detected in breast milk. Pediatr Infect Dis J. 2008; 27(6): 557-8
139. Bianchi DW, Flint AF, Pizzimenti MF, Knoll JH, Latt SA. Isolation of fetal DNA from nucleated erythrocytes in maternal blood. Proc Natl Acad Sci USA. 1990; 87(9): 3279-83
140. Mueller UW, Hawes CS, Wright AE, Petropoulos A, DeBoni E, Firgaira FA, et al. Isolation of fetal trophoblast cells from peripheral blood of pregnant women. Lancet. 1990(8709); 336: 197-200
141. Ganshirt D, Garritsen H, Miny P, Holzgreve W. Fetal cells in maternal circulation throughout gestation. Lancet. 1994; 343(8904): 1038-9
142. Vernochet C, Caucheteux SM, Kanellopoulos-Langevin C. Bi-directional cell trafficking between mother and fetus in mouse placenta. Placenta. 2007; 28(7): 639-49
143. Nijagal A, Wegorzewska M, Jarvis E, Le T, Tang Q, MacKenzie TC. Maternal T cells limit engraftment after in utero hematopoietic cell transplantation in mice. J Clin Invest. 2011; 121(2): 582-92
144. Tuboly S, Bernath S, Glavits R, Kovacs A, Megyeri Z. Intestinal absorption of colostral lymphocytes in newborn lambs and their role in the development of immune status. Acta Vet Hung. 1995; 43(1): 105-15
145. Tuboly S, Bernath S. Intestinal absorption of colostral lymphoid cells in newborn animals. Adv Exp Med Biol. 2002; 503: 107-14
146. Amir LH, Garland SM, Dennerstein L, Farish SJ. Candida albicans: is it associated with nipple pain in lactating women? Gynecol Obstet Invest. 1996; 41(1): 30-4
Copyright © 2013- by Cory H. E. R. & C. Inc. All Rights Reserved.