www.fimpolgy.com/JTF
The Journal of Theoretical Fimpology. Volume 1, Issue 4: e-20111024-1-4-8. December 22, 2013 (www.fimpology.com)
The Universal Pattern of Evolutionary Entities and Its Circulatory Ladder-Like Pyramid Feature
Shu-dong Yin
ORCID ID: https://orcid.org/0009-0005-8661-6889
Cory H. E. R. & C. Inc., Burnaby, British Columbia, Canada
During the past decades, the newly discovered sub-cellular and cellular entities have uncovered the insufficiency of the traditional Darwinians' Universal Tree of Life (TOL); and accumulating evidences have indicated that genetic alteration in both prokaryotes and eukaryotes was not only endogenous as what neo-Darwinians insisted on, but also exogenous as what Lamarckians claimed, which is challenging the Modern Synthesis-our modern knowledge on evolution as the traditional theoretical cornerstone of many disciplines in the life sciences including biology, physiology and medical science. Based on the advantages in multi-disciplines, the author proposes a novel theoretical model, called the Universal Pattern of Evolutionary Entities (UPOEE) to complement the inadequacies in TOL and the traditional definition of life. UPOEE reflects the path of evolution from the simple to the complex, from lower levels to higher levels, and from inorganic entities to living organisms. Moreover, in the UPOEE model, there are also several newly proposed concepts: (i) the Last Universal Inorganic Molecular Ancestor (LUIMA), and the Last Universal Organic Molecular Ancestor (LUOMA), and the Last Universal Sub-Cellular Ancestor (LUSCA) for illustrating the three important starting lines for demarcating evolution of molecular and sub-cellular entities from lower levels to higher levels in the UPOEE model, (ii) evoccasion, evolvamity and evoclash for elucidating the opportunity and interaction among evolutionary entities at the same or different evolutionary levels, and (iii) the 'circulatory ladder-like pyramid' pattern, a facet of UPOEE beside known 'tree-like', 'web-like', and 'goods-like' features.
Key words: tree of life; the Universal Pattern of Evolutionary Entities (UPOEE); evoccasion; evolvamity; evoclash
The concept of the tree of life (TOL) reflects the gradational characteristic of evolution, which was initially based on studies on the extant natural macro organisms, such as plants and animals, and the extinct macro organisms left in fossils.[1-5] However, the lowest starting point of evolution was unclear in the 19th century.[6] Later after entering the 20th century, the evolutionary root of TOL was excavated into the cellular level; and furthermore, the term 'procaryote' was coined for a cell without a nucleus and 'eucaryotes' for those having nuclei.[6, 7] Structurally and functionally, prokaryotic cells were simpler than eukaryotic cells of plants and animals. From the evolutionary perspective, prokaryotic cells should emerge earlier than eukaryotic cells and this notion has been supported by fossil evidences.[1-3, 8] As the only dominant unicellular life form before eukaryotes, prokaryotes were superior to the polyphasic entities of the sub-cellular evolutionary level, such as viruses, phages and vesicle-like entities.
As our excavating unknowns become deeper and vaster, newly discovered sub-cellular and molecular entities have no place in the traditional Darwinians' universal tree of life;[9] and accumulating evidences have indicated that genetic alteration in both prokaryotes and eukaryotes was not only endogenous as what neo-Darwinians insisted on, but also exogenous as what Lamarckians claimed, which is challenging the Modern Synthesis-our modern knowledge on evolution as the traditional theoretical cornerstone of many disciplines in the life sciences.[9] Moreover, the combination of Darwinian natural selection and Mendelian genetics, which resulted in the Modern Synthesis in the mid-20th century has been proved theoretically and empirically deficient in accounting for the question "how did and does evolution proceed?" [9]
'Tree-like', 'Web-like', and 'Goods-like' Features
Norman Pace pointed out that "while our understanding of the diversity and evolution of microbial life is advancing rapidly, our textbooks are not incorporating new information or concepts at nearly the same pace. One important consequence is that textbook explanations of life's organization and evolutionary history are badly outdated." [6] Upon many discoveries, such as viruses, plasmids, prions, transposable elements, and horizontal gene transfer (HGT) in the 20th century, the root of TOL has been excavating into the sub-life level. Especially, Archaea found by Woese has indicated that the root of TOL is not at the cellular level,[10] which led to recent theoretical explorations on the origin of cells,[11-21] and debates on the necessity and the universality of TOL,[5, 6, 22-29] while some new theoretical models were proposed as well. For example, Martin argued that "microbial genome evolution is not a treelike process because of lateral gene transfer and the endosymbiotic origins of organelles," "the tree of life is only one impediment to a better understanding of early evolution...we might not need a tree of life at all."[22] Logsdon claimed that "the prokaryotic tree of life is dead!"[5] McInerney and colleagues proposed 'the public goods hypothesis', in which, "nucleotide sequences (genes, promoters, exons, etc.) are simply seen as goods, passed from organism to organism through both vertical and horizontal transfer." [30] and moreover, Torday and Rehan revealed that the molecular homologies of parathyroid hormone-related protein (PTHrP), a peptide secreted by alveolar type II cells, were found to exist in the lung, adrenals, skin, kidney, gut, bone, and brain of extant land vertebrates.[31, 32] Kutschera wrote that "Darwin's view of a static, 'Animals and Plants-based Tree of Life' that does not take into account micro-organisms and endosymbiotic events, is outdated." And Kutschera proposed a tree-like 'symbiogenesis, natural selection, and dynamic Earth (synade)-model', in which symbiogenesis, (directional) natural selection, and the dynamic Earth were viewed "as three important 'driving forces' of organismic evolution".[24, 33, 34] Recently, O'Malley and Koonin concluded that "the irrefutable demonstration by phylogenomics that different genes in general have distinct evolutionary histories made obsolete the belief that a phylogenetic tree of a single universal gene such as rRNA or of several universal genes could represent the 'true' TOL." [23] And they suggested that the traditional Darwinian TOL may be 'replaced by new, probably web-like representations of genome evolution',[23] or may 'call for a radical revision (if not abandonment) of the Tree of Life concept'.[35] Moreover, some authors revealed another essential trait of evolutionary pattern but missed in TOL, which is a net or web-like characteristic.[23, 36-38]
In my opinion, although viruses have no place in the Darwinians' tree of cellular life, they play an important role in both pre-life and life evolution, and therefore, actually have unshakable places in the pre-cellular universal tree.[9] The pre-life's tree, as the extension of the root of tree of cellular life, encompasses all relevant inorganic and organic molecular entities. Here, we do not need to re-define "what is life or organism?" because that there are different definitions for it at different evolution levels and that while excavating the evolutionary root deeper and deeper,[8] the only thing we should do is to put newly found evolutionary pieces in a suitable place, not to replace or change the old parts in the evolution puzzle. Therefore, I propose a new tree named 'the universal tree of evolutionary entities' to combine the Darwinians' tree of cellular life with the pre-cellular universal tree, in which the relevant inorganic and organic molecular entities, viruses, bacteria, archaea and eukaryotes have their own specific evolutionary places, and constitute different evolutionary platforms. The study on evolution should be a specific interest not only in the life sciences, but also in physics and chemistry.
The traditional TOL is not an obstacle, but a supplement for understanding the Universal Pattern of Evolutionary Entities (UPOEE); and moreover, it has its historical and real life in macro-biological world because of its root at the cellular level, and therefore is an indispensable part of UPOEE. 'Web-like' and 'goods-like' patterns of prelife evolutionary entities could be understood as the important development and extension of TOL, the evolutionary background and mechanism of TOL, and a new piece in the UPOEE puzzle. I agree to the view that prokaryotic microorganisms are the evolutionary 'background organisms' of eukaryotic multi-cellular macroorganisms;[24] and I further propose a concept of 'Evolutionary Background Entities, EBE', which refers to those entities of the lower evolutionary levels. According to this definition, EBE of eukaryotic multi-cellular macroorganisms embrace not only prokaryotic entities, but also include all prelife evolutionary entities, such as viruses, phages, genes, and other relevant organic and inorganic entities.
Evolutionary Gradation of UPOEE
As the consequence of evolution, the emergence of diversification and gradation of evolutionary entities at the population level enabled them to have obtained novel traits or capabilities for their survival. In the UPOEE model (Figure 1), the whole evolution can be gradated by the following different ways: (i) the Mono-Phase Period and the Poly-Phase Period are proposed according to the structural phase of evolutionary entities. The Mono-Phase Period refers to the ancient evolutionary period when monomers and polymers existed independently as monophasic entities, such as inorganic elements and compounds and organic complex without any envelope-like surrounding or coating structures; and therefore, the environment of an evolutionary entity in the Mono-Phase Period should be more dynamic and changeable than that in the Poly-Phase Period. In contrast, the Poly-Phase Period refers to the ancient evolutionary period when monophasic entities were enclosed by membrane or coating structure
to form evolved polyphasers, such as sub-cellular and cellular entities, which, however, evolved a relatively stable interior environment; (ii) the Molecular Stage, the Sub-Cellular Stage, the Unicellular Stage, and the Multi-Cellular Stage are based on the complexity of evolutionary entities. Evolutionary entities of the Molecular Stage match that of the Mono-Phase Period. The remaining stages belong to the Poly-Phase Period; and (iii) according to the extant entities, the whole evolutionary entities could be distributed into different levels via the order from the top (the most complicate in structure) to the bottom (the simplest in structure) as the following: the level of multi-cellular macroorganisms, the level of unicellular eukaryotes, the level of unicellular prokaryotes, the level of sub-cellular entities, the level of organic entities, and the level of inorganic entities (Figure 1).
Moreover, there is a necessarily evolutionary course from an individual to a group or population for any given evolutionary entity at the relevant evolutionary level to form a species at the population level. Evolutionary entities of each evolutionary level had/have three common characteristics: (a) relevant evolutionary entities at the same evolutionary level were/are similar in the evolutionary degree of structural complexity from the evolutionary perspective; (b) despite of being at the same evolutionary level, they were/are diverse in structural composition and functional behavior, for which, they could/can be divided as different species at the population level and separated as the unique individuals at the individual level; and (c) evolutionary entities of any given higher evolutionary level were/are composed of evolutionary entities at the lower evolutionary levels (Figure 2).
The UPOEE model could also be dissected into several levels for the distribution of different kinds of evolutionary entities (Figure 1). Moreover, evolutionary entities at the same evolutionary level varied morphologically, structurally and functionally. In animals, evolutionary entities at the macroorganism individual level and the macroorganism population level have different evolutiological significance. In human beings, the macroorganism population level could be further divided into several sub-levels, such as the group level, the tribe level, the national level, and the international level. An entity at the national level is called a 'country' or a 'nation', and at the international level, it is an international or world organization.
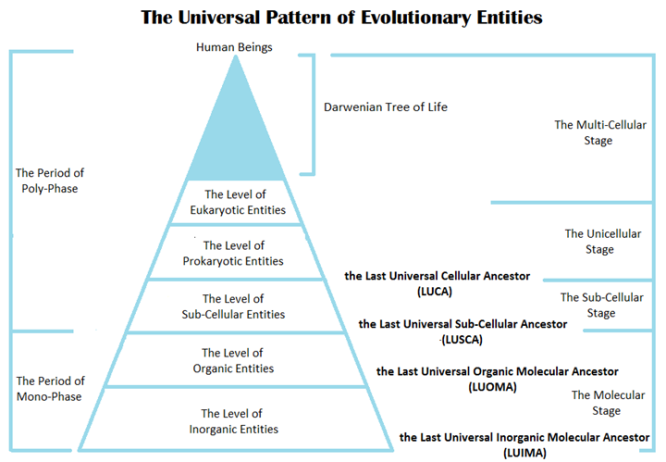
Figure 1: In the model of the Universal Pattern of Evolutionary Entities, the evolution order from the simple to the complex is illustrated from three aspects as the following: (1) the period of monophase to the period of polyphase, (2) the molecular stage to the sub-cellular stage, then the unicellular stage, and then to the multi-cellular stage, and (3) the level of inorganic entities to the level of organic entities, then the level of sub-cellular entities, then the level of prokaryotic entities, and then to the level of eukaryotic entities.
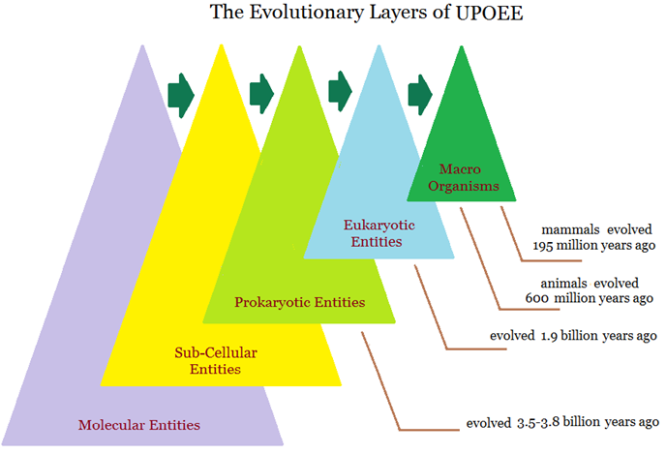
Figure 2: The evolutionary milestones or the evolutionary layers are illustrated in the model of the Universal Pattern of Evolutionary Entities. For a given evolutionary entity, those evolutionary entities of self at the lower levels comprise its biological and non-biological background and all those non-self evolutionary entities may constitute its ecological and evolutiological environment.
The Evolutionary Levels at the Molecular Stage
Evolutionary entities at the inorganic molecule level include H, O, N, P, S, Ca, Na, K, Fe, Mg, Si, Ni, H2, H2O, NH3, H2S, and SO2; and CO, CO2, CH4, amino acids, nucleotides, lipids and carbohydrates etc. are the exemplar of evolutionary entities at the organic molecule level. The members at these two levels cover a wide range of molecular entities including abiotic elements, abiotic inorganic and organic compounds, which were initially concerned by chemistry and physics, then by biochemistry.[39-48] In the UPOEE model, the Last Universal Organic Molecular Ancestor (LUOMA), the Last Universal Inorganic Molecular Ancestor (LUIMA) and the Last Universal Sub-Cellular Ancestor (LUSCA) are proposed as the three important starting lines during the molecular and sub-cellular stages of prelife (or prebiotic) evolution (Figure 1). Therefore, theoretically, LUIMA, LUOMA and LUSCA are the reasonable extension of the concept 'the last universal common ancestor of viruses, LUCAV', which was proposed for presenting the ancient viral entities at the sub-cellular level of prelife evolution.[13]
Monomers to Polymers
Monomers in this paper refer to chemical elements. A polymer is a compound consisted by similar and/or different monomers linked by various bonds; and a monomer is the building block of a polymer. Proteins, carbohydrates, DNA and RNA are polymers, and emerged as monophasic entities during the Mono-Phase Period of evolution. Monophasers are those evolutionary entities including monomers and polymers that lack any enclosing structures. Evolution from monomers to polymers to form organic macromolecules with biochemical functions of genetic information transfer and catalysis was one of the premises for the origin of primary cellular structures.[49-62] More and more research attention from the evolution perspective has been paid to the prelife monophasers between LUOMA and LUIMA,[63-75] which has revealed some new features of evolutionary molecular entities; for instance, studies on protein conformation-based inheritance [76-79] has clearly indicated that at the molecular level, besides polyribonucleotides such as RNA and DNA, some specific oligopeptides can also play a role as hereditary information carriers.[9] RNA, a polyribonucleotide can play a role as catalytic enzymes besides its well-known role as templates for replication and read-out of genetic information.[9, 80, 81]
Monophasers to Polyphasers
Any evolutionary entities that are enclosed by coating structures which are composed mainly by peptides, carbohydrates, or lipids, are polyphasic entities, and could be called polyphasers, which include viruses, phages, microvesicles, prokaryotic cells and eukaryotic cells. Monomers, polymers, monophasers and polyphasers reflect different responding evolutionary layers, and also represent different milestones in the path of evolution. Furthermore, if there shouldn't be some organic polymers to evolve from monophase to polyphase, the origin of cellular life would not have been realized;[82] and this important evolutionary transform was dependent on the emergence of other two polymers, called 'membrane' and cytoplasm' as the critical premise for the emergence of the most primitive cells-like polyphasers.
The most remarkable difference between monophasers and polyphasers is that a polyphaser has a relative stable interior environment which monophasers lack even though both monophasers and polyphasers have their own external environments which directly surround them at the individual level. As the consequence of the transition from monophase to polyphase, previous monophasic entities evolved to a higher evolutionary level, enabling a group of the previous monophasic entities to have a relatively stable internal environment where more complicated and efficient physical and chemical interactions could occur. In addition, according to the composition of core polymers in coating structures, three major classes of extant evolutionary polyphasic entities may be divided as the following: (i) peptide-shell-coated entities, such as viruses and phages, (ii) carbohydrate-shell-coated entities, such as cell wall-enveloped plant cells, and (iii) plasma membrane-coated entities, such as animal eukaryotic cells. The importance of plasma membrane from the evolution perspective can be elucidated further from the research made by Randy Schekman, Thomas Sudhof and James Rothman, who were awarded the Nobel Prize in Physiology or Medicine in 2013.[83]
The Evolutionary Levels at the Sub-cellular and Cellular Stages
Viruses, phages, cyto-organelles, and exosomes are the extant representative of evolutionary entities at the sub-cellular level; bacteria, archaea, fungi are the extant exemplars of evolutionary entities at the unicellular eukaryotic and prokaryotic levels; and evolutionary entities at the multi-cellular macroorganism level consist of all plants and animals in the traditional Darwinians' Universal TOL. Gould pointed out that the first five-sixths of cellular life history were almost exclusively unicellular.[4] Clearly, the unicellular prokaryotic level was a rather long period on the time scale of evolution, during which, certainly there might be numerous transitional prokaryotic cellular forms that were extinct finally, which resulted in only extant bacteria and archaea we can see today.
New Capabilities-as the Consequence of Evolution for Survival Varied at the Different Evolutionary Levels
Prelife and living entities obtained novel traits or functions through evolution for their survival; and therefore, at different evolutionary levels, corresponding evolutionary entities display different capabilities, functions and behaviors. Although the definition of life is diverse, blurred and undetermined, the functions of evolutionary entities at different evolutionary levels are truly distinctive. For example, animals at the individual level of macroorganism have evolved several unique functions, capabilities, and behaviors, such as fighting, foraging, sexual mating, vocal signals and social learning, which are impossible for the evolutionary entities of the cellular level to display.
Evolutionary entities at the molecular level cannot demonstrate the functions, capabilities or behaviors displayed by the entities at the cellular level, such as cellular migration, responding to an internal and external cellular stimulus, cellular growth and division despite of facts that these cellular behaviors are actually the consequence of hundred thousands of molecular interactions at the molecular level that occurred within cells. A human individual's behaviors, such as speaking, watching, walking, singing and thinking, are a few exemplars of the functions that we can display at the macroorganism individual level, which, quite well-known today, are the comprehensive consequence of a complex cooperation of numerous cells and organ systems in our bodies. The population level of some animal species is also called the social level. Several social behaviors, such as foraging, reproduction and social learning have been observed naturally or studied in some animal species including ants, birds, fishes, rats, dogs, and monkeys.[84-95]
As the most advantaged mammal species in the evolution process, modern human beings currently stand on the top of UPOEE. However, humans' unconquerable today is the evolutionary consequence of the past hundred thousand years as the inferior species in the timescale of evolution not only when the ancestors of hominids were in the ancient mammal period, but also when that evolved to the primate level. Since humans entered the era of modern civilization, newly evolved social behaviors, such as cultures and arts, economical, educational, political, legal and healthcare systems, military systems and warfare, religious activities, science and technology, and governments and international organizations have emerged and developed further as the unique symbols of humans at the social level. Therefore, it has been gradually accepted that the social feature of humans and non-human animals is deeply rooting in their biological nature. Unfortunately, such recognition had been neglected or rejected for a long time; and even today, the recognition that social system is the extension of biological evolution and that social problems of human beings are rooted in their biological nature is still a challenge to some sociologists, politicians, religionists, and even some biologists. One of the major reasons for their opposing opinion may be the extant picture of the whole evolution after the past several billion years is extremely fragmentary, from which, only micro-, macro-organisms dissimilar to each other is what we can see and moreover, tremendously diverse definitions of life and species are what we therefore formulated.[8]
Significance of the UPOEE model
The evolutionary mechanism of macro-organisms has been accounted for by the Modern Synthesis theory mainly from the molecular and macroorganism population perspective, in which, the molecular alterations of random mutation, genetic recombination and genetic drift are believed by neo-Darwinians to be the only source of genetic variation,[96-98] while interpreting macroorganism evolution for the notion of reductionism that "everything is reduced to the sum of the parts; and if we know the parts we can understand the whole." [98] Such DNA-centered neo-Darwinian reductionism was accepted as a mainstream theory in the life sciences during the past century. However, during the past decade, such reductionism understanding has been uncovered to suffer a congenital deficiency because much significance of evolutionary levels has been ignored.
The fimpological view is that knowing structurally the parts, or even all parts, of a thing cannot equal to understanding functionally the whole of it. In other words, a coordinate function or behavior of an evolutionary entity that is composed by entities at the lower multi-evolutionary levels cannot accurately be deciphered by focusing only on the entities of the lower evolutionary levels and without systemically considering their unique interactions at the higher level; and furthermore, a given evolutionary entity that evolved from a lower level to a higher level actually is always accompanied by obtaining novel functions or traits for its adaptation in a new environment, or its correspondence to some changes in the previously adapted environment in order to survive. For example, a cell can be divided into two parts: cytoplasm and nucleus at the sub-cellular level; and cell heredity or cell duplication, as an evolved function displayed at the cellular level that reflects the consequence of the interaction between cytoplasm and nucleus. In fact, the importance of cytoplasmic-nuclear interaction at the sub-cellular and molecular levels has been revealed via many empirical studies.[99-107]
Therefore, the cytoplasmic role in inheritance cannot be neglected while we talk about biological heritage at the cellular level. Moreover, if we discuss biological heritage of mammals at the macroorganism individual level, female-relayed intact cell inheritance should not be overlooked, which has been discussed in recent theoretical papers.[108-110] I argued that although evolution and nature selection occurs at all evolutionary levels, the mechanisms for evolution of macro-organisms at the individual level should not only proceed at the molecular level, but also operate at both the unicellular and the multi-cellular levels and the sub-cellular level. If we set up cells as the starting point of life, the tendency of evolution on Earth could be divided into two distinct parts: (i) the evolutionary age for pre-biological life, during which, the earlier the evolutionary entities evolved, the more remarkable the physical and chemical features would display. And (ii) the evolutionary age for cellular life, during which, the latter the evolutionary entities evolved, the more notable the social features would exhibit. Moreover, during evolution, initial physical and chemical nature of evolutionary entities has been sheltered by brilliant biological features and then, biological nature of them is shadowed further by social characters. However, those sheltered physical, chemical and biological characters, in fact, have never lost or disappeared, and occasionally demonstrated their existence as the forms of various human super-powers, most of which may have been attributed to 'mysterious natural phenomena'. Recently, the emergence of a novel disciplinary-Mechanobiology, which studies the essential roles of physical factors playing via the process of mechanotransduction from the molecular, nuclear, cellular levels to the tissue level [111, 112] has initiated a new exploration for the physical nature of macroorganism individuals.
Evolutionary entities at the eukaryotic cell level consist of unicellular eukaryotic cells and multi-cellular eukaryotic cells. Multi-cellular eukaryotic cells have a unique characteristic which is that they need an embryonic period or a series of growth and developmental stages to complete their transformation from a single fertilized cell to a multi-cellular macroorganism individual. Plants and animals are the best embodiment of such unique characteristic. In fimpological view, each individual of evolutionary entities of a given evolutionary level should be embodied in a corresponding collection or a species at the population level; and evolutionary entities at the higher evolutionary levels evolved from and are composed of entities that belong to the lower evolutionary levels. For example, a human body at the macroorganism individual level is a single evolutionary entity; however, at the cellular level, an adult human body contains more than 10(13) host eukaryotic cells and 10(14-15) prokaryotic cells;[108, 113-115] and there should be a much higher number of evolutionary entities at the lower levels, such as the sub-cellular and molecular levels. According to the Modern Synthesis's notion that evolution occurs at a species and not at macroorganism individuals of a species,[116] a paradoxical question arises as the following: a human body at the population level would not be considered to be able to evolve; and however, at the cellular level, there are different host eukaryotic cellular and prokaryotic cellular species or lineages in a human body and can they evolve? Similar questions could arise at the sub-cellular level and the molecular level. In fact, three decades ago, Ernst Mayr already admitted that "how does a given evolutionary principle or phenomenon actually operate in an individual case?" was a question that the Modern Synthesis had not answered yet.[117] Clearly, from the fimpological perspective, the Modern Synthesis neglected genetic and non-genetic variation-induced evolution occurred at the sub-cellular and cellular levels.
Relationship and Interaction among Different Evolutionary Levels: Evolvamity, and Evoclash
In the fimpological view, evolution happens all the time at every evolutionary level,[9] for which, it's necessary to develop some novel concepts to describe the relationship and interaction of evolutionary entities among different evolutionary levels. In this paper, I propose two novel terms: evolvamity and evoclash, as the expansion of the term 'evolvability'. 'Evolvamity' refers to the harmony of evolutionary entities between different evolutionary levels or within the same evolutionary level; and 'evoclash' refers to the discord of evolutionary entities between different evolutionary levels or within the same evolutionary level (Figure 1 and 2).
In fact, the interaction between different evolutionary levels has already been mentioned before by a few researchers. For instance, Michod, Nedelcu and Roze pointed out that there are often conflicts or disagreements between the higher-level and the lower level;[118] and Paenke, Jin and Branke argued that the dynamic environment is a decisive factor for the balance between the individual level and the population level.[119] The cooperation of different nations, such as the Central Powers and the Alies were the example of evolvamity; in contrast, the first and second world wars were the example of evoclash at the social level. Moreover, the phenomenon of the six models of pregnancy-associated eukaryotic cell transmission between the fetus and mother, and the phenomenon of infant fetal cells co-existing normally within maternal organs and tissues [109] are the exemplar of evolvamity between the cellular level and the individual level. In contrast, diseases of plants, humans, and non-human animals may be the case of evoclash between pathogenic evolutionary entities of the individual level, the sub-cellular level and the molecular level, and host evolutionary entities of the individual level, the cellular level, the sub-cellular level and the molecular level.
Obscure viral infections are the consequence of evoclash between the pathogenic viral sub-cellular level and the host molecular and cellular levels, but not that between the viral sub-cellular level and the host individual; however, if viral infections develop into clinical diseases, their evoclash correspondingly involve in the viral sub-cellular level and the host molecular, cellular and individual levels. Organic phosphorus herbicide-induced clinical diseases reflect evoclash between the toxic molecular level and the hominal host molecular, cellular and individual levels. Homicidal death reflects the different consequences of entities' interactions at or between different evolutionary levels: (i) the consequence of evolvamity among murders' molecular, cellular and individual levels for operating homicidal idea and act, (ii) the consequence of evoclash between murders' individual level and victims' individual level; and (iii) the legal penalty faced by murders exemplifies the evoclash between murders' individual level and the social level of the human society.
Evoccasion-Unequal Chance for All Evolutionary Entities at the Population Level
It is worth to point out that not all individual members of a species can equally catch an evolutionary opportunity. In other words, the opportunity of encountering evolution or evoccasion is unequal to each member of a species. For example, destructive volcanic eruptions and earthquakes were usually regional geological events, which caused the previous micro- and macro-environmental conditions of the affected area to alter, and resulted in an active-environmental-alteration-driven evolution process to befall the microorganisms and macroorganisms including plants and animals within that region or area, but usually did not affect the microorganisms and macroorganisms of the same species which were out of the region; and therefore, a new term 'evoccasion' is proposed for such inequality.
The capacity of entities to evolve has been called evolutionary adaptability, or evolvability, which has been widely discussed at the molecular, cellular, and individual levels;[120-126] and however, what evolvability concerns is focused on the internal opportunity and mechanism of evolutionary success for those evolutionary entities under nature selection. In contrast, evoccasion refers to the external opportunities of evolution that will initiate a process of nature selection acting on targeting evolutionary entities. Evoccasional variation may be an initial environmental or external reason for diversity which has occurred in a widely dispersed species.
Opportunity and spatiality are the two important characters of evoccasion, which means that (i) the occurrence of environmental alterations, or events including volcano eruptions, earthquakes, desert formation, and collisions of asteroids and comets with Earth usually last a certain period with the variety from seconds to years, and befall the micro- and macroorganism entities within a limited space and area; (ii) the majority of evoccasional events occurred only one time in the timescale of evolution, which means that not all of the individual members of a species at an evolutionary level would have an equal opportunity to evolve to a higher evolutionary level; and (iii) the evolutionary consequences are usually uneasy to be reversed or to copy mainly because of that we still know a little about the history of evolution and that it's uneasy to simulate realistically a series of regionally and geologically evoccasional events.
The Circulation-Based Ladder-like Pyramid Model
Moreover, beside of 'tree-like', 'web-like', 'goods-like' and external environment-driven features, UPOEE should have another facet-'Circulation-Based Ladder-like Pyramid' pattern. In other words, there are several cyclic features in the UPOEE model. One of them is the cycle operating between lower levels and higher levels, among which, the carbon cycle is one and another is the cycle working between individuals and populations. The ladder-like pyramid pattern reflects the relationship of continuous evolutionary layers according to the time scale of evolution, in which, human beings on the top and inorganic entities on the bottom. The further discussion of the third circulatory feature of UPOEE will be later in other papers.
In the UPOEE model (Figure 1 and 2), the shape of 'Pyramid' means that the number of entities in the higher-levels is always smaller than that of entities in the lower levels from where the entities in the higher-levels evolved, which is the consequence of evoccasional inequality. For instance, at the sub-cellular level, viruses (bacteriophages) are the most abundant evolutionary bioentities on earth, with approximately 10(31) viral particles in the oceans;[127-131] at the unicellular level, the number of prokaryotes on Earth was estimated to be 10(30) cells;[132] and at the macroorganism population level, humans' population is 7.2x10(9) [133].
Summary
In this paper, the author proposes a novel theoretical model, called UPOEE to complement the inadequacies in TOL, in addition to several new concepts, such as LUIMA, LUOMA and LUSCA for illustrating the three important starting lines for demarcating evolution of molecular and sub-cellular entities from a lower level to a higher level in the UPOEE model, and evoccasion, evolvability, evolvamity and evoclash for elucidating the opportunity and interaction among evolutionary entities at the same or different evolutionary levels, as well as the 'circulatory ladder-like pyramid' pattern, a facet of UPOEE beside known 'tree-like', 'web-like' and 'goods-like' characters.
Abbreviations
TOL: the Tree of Life;
UPOEE: the Universal Pattern of Evolutionary Entities;
EBE: Evolutionary Background Entities;
LUIMA: the Last Universal Inorganic Molecular Ancestor;
LUOMA: the Last Universal Organic Molecular Ancestor;
LUSCA: the Last Universal Sub-Cellular Ancestor;
LUCA: the Last Universal Cellular Ancestor;
LUCAV: the Last Universal Common Ancestor of Viruses.
References
1. Knoll AH. The early evolution of eukaryotes: a geological perspective. Science. 1992; 256(5057): 622-7
2. de Duve C. The beginnings of life on Earth. Am Sci. 1995; 83(5): 428-37 (Available from: http://www.americanscientist.org/issues/num2/the-beginnings-of-life-on-earth/1 Accessed 12-22-2013)
3. Schopf JW. Are the oldest fossils cyanobacteria? Evolution of microbial life: fifty-fourth Symposium of the Society for General Microbiology held at the University of Warwick, edited by Roberts DM, Sharp P, Alderson G, Collins MA. Pp.23-61. The Press Syndicate of the University of Cambridge, New York. USA. 1996
4. Gould JS. The evolution of life. Evolution! Facts and Fallacies. Ed. by J. William Schopf. Pp.1-13. Academic Press, California, USA. 1999
5. Bapteste E, O'Malley MA, Beiko RG, Ereshefsky M, Gogarten JP, Franklin-Hall L, et al. Prokaryotic evolution and the tree of life are two different things. Biol Direct. 2009; 4: 34
6. Pace NR. The molecular tree of life changes how we see, teach microbial diversity. Microbe. 2008; 3(1): 15-20
7. Doolittle WF, Zhaxybayeva O. On the origin of prokaryotic species. Genome Res. 2009; 19(5): 744-56
8. Yin S-d. To define life and species as to demarcate the International Date Line. The Journal of Theoretical Fimpology. 2013; 1(4): e-20111129-1-4-7. Available from: http://www.fimpology.com
9. Yin S-d. A fimpological view: The future synthesis of biology, ecology, and evolutiology. The Journal of Theoretical Fimpology. 2013; 1(3): e-20080225-1-3-5. Available from: http://www.fimpology.com
10. Doolittle WF. Phylogenetic classification and the universal tree. Science. 1999; 284(5423): 2124-9
11. Margulis L. Archaeal-eubacterial mergers in the origin of Eukarya: phylogenetic classification of life. Proc Natl Acad Sci USA. 1996; 93(3): 1071-6
12. Woese CR. On the evolution of cells. Proc Natl Acad Sci USA. 2002; 99(13): 8742-7
13. Koonin EV, Senkevich TG, Dolja VV. The ancient Virus World and evolution of cells. Biol Direct. 2006; 1: 29
14. Rocheleau T, Rasmussen S, Nielsen PE, Jacobi MN, Ziock H. Emergence of protocellular growth laws. Philos Trans R Soc Lond B Biol Sci. 2007; 362(1486): 1841-5
15. Cox CJ, Foster PG, Hirt RP, Harris SR, Embley TM. The archaebacterial origin of eukaryotes. Proc Natl Acad Sci USA. 2008; 105(51): 20356-61
16. Yutin N, Makarova KS, Mekhedov SL, Wolf YI, Koonin EV. The deep Archaeal roots of eukaryotes. Mol Biol Evol. 2008; 25(8):1619-30
17. Archibald JM. The eocyte hypothesis and the origin of eukaryotic cells. Proc Natl Acad Sci USA. 2008; 105(51): 20049-50
18. Loakes D, Holliger P. Darwinian chemistry: towards the synthesis of a simple cell. Mol Biosyst. 2009; 5(7): 686-94
19. Cavalier-Smith T. Origin of the cell nucleus, mitosis and sex: roles of intracellular coevolution. Biol Direct. 2010; 5: 7
20. Schrum JP, Zhu TF, Szostak JW. The origins of cellular life. Cold Spring Harb Perspect Biol. 2010 May 19 [PubMed]
21. Cavalier-Smith T. Deep phylogeny, ancestral groups and the four ages of life. Phil Trans R Soc B. 2010; 365(1537): 111-32
22. Martin WF. Early evolution without a tree of life. Biol Direct. 2011; 6: 36
23. O'Malley MA, Koonin EV. How stands the Tree of Life a century and a half after The Origin? Biol Direct. 2011; 6: 32
24. Kutschera U. From the scala naturae to the symbiogenetic and dynamic tree of life. Biol Direct. 2011; 6: 33
25. Theobald DL. A formal test of the theory of universal common ancestry. Nature. 2010; 465(7295): 219-22
26. Koonin EV, Wolf YI. The fundamental units, processes and patterns of evolution, and the tree of life conundrum. Biol Direct. 2009; 4: 33
27. McInerney JO, Cotton JA, Pisani D. The prokaryotic tree of life: Past, present ... and future? Trends Ecol Evol. 2008; 23(5): 276-81
28. Philippe H, Forterre P. The rooting of the universal tree of life is not reliable. J Mol Evol. 1999; 49(4): 509-23
29. Baldauf SL, Palmer JD, Doolittle WF. The root of the universal tree and the origin of eukaryotes based on elongation factor phylogeny. Proc Natl Acad Sci USA. 1996; 93(15): 7749-54
30. McInerney JO, Pisani D, Bapteste E, O'Connell MJ. The public goods hypothesis for the evolution of life on Earth. Biol Direct. 2011; 6: 41
31. Torday JS, Rehan VK. Exploiting cellular-developmental evolution as the scientific basis for preventive medicine. Med Hypotheses. 2009; 72(5): 596-602
32. Torday JS. Evolution and Cell Physiology. 1. Cell signaling is all of biology. Am J Physiol Cell Physiol. 2013; 305(7): C682-9
33. Kutschera U. Plant-associated methylobacteria as co-evolved phytosymbionts. A hypothesis. Plant Signal Behav. 2007; 2(2): 74-8
34. Whitman WB, Coleman DC, Wiebe WJ. Prokaryotes: the unseen majority. Proc Natl Acad Sci USA. 1998; 95(12): 6578-83
35. Koonin EV, Wolf YI. Evolution of microbes and viruses: a paradigm shift in evolutionary biology? Front Cell Infect Microbiol. 2012; 2: 119
36. Olendzenski L, Gogarten JP. Evolution of genes and organisms: the tree/web of life in light of horizontal gene transfer. Ann NY Acad Sci. 2009;1178: 137-45
37. Dagan T, Martin W. Getting a better picture of microbial evolution en route to a network of genomes. Phil Trans R Soc B. 2009; 364(1527): 2187-96
38. Puigbo P, Wolf YI, Koonin EV. The tree and net components of prokaryote evolution. Genome Biol Evol. 2010; 2: 745-56
39. Hazen RM, Sverjensky DA. Mineral surfaces, geochemical complexities, and the origins of life. Cold Spring Harb Perspect Biol. 2010; 2(5): a002162
40. Van Mooy BA, Fredricks HF, Pedler BE, Dyhrman ST, Karl DM, Koblízek M, et al. Phytoplankton in the ocean use non-phosphorus lipids in response to phosphorus scarcity. Nature. 2009; 458(7234): 69-72
41. Forterre P, Gribaldo S. The origin of modern terrestrial life. HFSP J. 2007; 1(3): 156-68
42. Jortner J. Conditions for the emergence of life on the early Earth: summary and reflections. Philos Trans R Soc Lond B Biol Sci. 2006; 361(1474): 1877-91
43. Deamer D, Singaram S, Rajamani S, Kompanichenko V, Guggenheim S. Self-assembly processes in the prebiotic environment. Phil Trans R Soc B. 2006; 361(1474): 1809-18
44. Morowitz HJ, Kostelnik JD, Yang J, Cody GD. The origins of intermediary metabolism. Proc Natl Acad Sci. 2000; 97(14): 7704-8
45. Kornberg A, Rao NN, Ault-Riche D. Inorganic polyphosphate: A molecule of many functions. Annu Rev Biochem. 1999; 68: 89-125
46. Thaler DS, Liu S, Tombline G. Extending the chemistry that supports genetic information transfer in vivo: phosphorothioate DNA, phosphorothioate RNA, 2'-O-methyl RNA, and methylphosphonate DNA. Proc Natl Acad Sci USA. 1996; 93(3): 1352-6
47. Westheimer FH. Why nature chose phosphates. Science. 1987; 235(4793): 1173-8
48. Wachtershauser G. Evolution of the first metabolic cycles. Proc Natl Acad Sci USA. 1990; 87(1): 200-4
49. Ferris JP, Hill AR Jr, Liu R, Orgel LE. Synthesis of long prebiotic oligomers on mineral surfaces. Nature. 1996; 381(6577): 59-61
50. Bartel DP, Unrau PJ. Constructing an RNA world. Trends Cell Biol. 1999; 9(12): M9-13
51. Franchi M, Ferris JP, Gallori E. Cations as mediators of the adsorption of nucleic acids on clay surfaces in prebiotic environments. Orig Life Evol Biosph. 2003; 33(1): 1-16
52. Bada JL, Lazcano A. Origin of life: Some like it hot, but not the first biomolecules. Science. 2002; 296(5575): 1982-3
53. Orgel LE. Some consequences of the RNA world hypothesis. Orig Life Evol Biosph. 2003; 33(2): 211-8
54. Ertem G. Montmorillonite, oligonucleotides, RNA and origin of life. Orig Life Evol Biosph. 2004; 34(6): 549-70
55. Franchi M, Gallori E. A surface-mediated origin of the RNA world: biogenic activities of clay-adsorbed RNA molecules. Gene. 2005; 346: 205-14
56. Ferris JP. Montmorillonite-catalysed formation of RNA oligomers: the possible role of catalysis in the origins of life. Philos Trans R Soc Lond B Biol Sci. 2006; 361(1474): 1777-86
57. Cleaves HJ 2nd. The origin of the biologically coded amino acids. J Theor Biol. 2010; 263(4): 490-8
58. Nowak MA, Ohtsuki H. Prevolutionary dynamics and the origin of evolution. Proc Natl Acad Sci USA. 2008; 105(39): 14924-7
59. Johnson AP, Cleaves HJ, Dworkin JP, Glavin DP, Lazcano A, Bada JL. The Miller volcanic spark discharge experiment. Science. 2008; 322(5900): 404
60. Bryant JA, Aves SJ. Initiation of DNA replication: functional and evolutionary aspects. Ann Bot. 2011; 107(7): 1119-26
61. Parker ET, Cleaves HJ, Dworkin JP, Glavin DP, Callahan M, Aubrey A, et al. Primordial synthesis of amines and amino acids in a 1958 Miller H2S-rich spark discharge experiment. Proc Natl Acad Sci USA. 2011; 108(14): 5526-31
62. Gomolplitinant KM, Saier MH Jr. Evolution of the oligopeptide transporter family. J Membr Biol. 2011; 240(2): 89-110
63. Keefe AD, Miller SL, McDonald G, Bada J. Investigation of the prebiotic synthesis of amino acids and RNA bases from CO2 using FeS/H2S as a reducing agent. Proc Natl Acad Sci USA. 1995; 92(25):11904-6
64. Heinen W, Lauwers AM. Organic sulfur compounds resulting from the interaction of iron sulfide, hydrogen sulfide and carbon dioxide in an anaerobic aqueous environment. Orig Life Evol Biosph. 1996; 26(2): 131-50
65. Brandes JA, Boctor NZ, Cody GD, Cooper BA, Hazen RM, Yoder HS Jr. Abiotic nitrogen reduction on the early Earth. Nature. 1998; 395(6700): 365-7
66. Summers DP. Ammonia formation by the reduction of nitrite/nitrate by FeS: ammonia formation under acidic conditions. Orig Life Evol Biosph. 2005; 35(4): 299-312
67. Sleep NH, Zahnle K, Neuhoff PS. Initiation of clement surface conditions on the earliest Earth. Proc Natl Acad Sci USA. 2001; 98(7): 3666-72
68. Blencowe DK, Morby AP. Zn(II) metabolism in prokaryotes. FEMS Microbiol Rev. 2003; 27(2-3): 291-311
69. Lunine JI. Physical conditions on the early Earth. Philos Trans R Soc Lond B Biol Sci. 2006; 361(1474): 1721-31
70. Bernstein M. Prebiotic materials from on and off the early Earth. Philos Trans R Soc Lond B Biol Sci. 2006; 361(1474): 1689-700
71. Smirnov A, Hausner D, Laffers R, Strongin DR, Schoonen MA. Abiotic ammonium formation in the presence of Ni-Fe metals and alloys and its implications for the Hadean nitrogen cycle. Geochem Trans. 2008; 9: 5
72. Brandes JA, Hazen RM, Yoder HS Jr. Inorganic nitrogen reduction and stability under simulated hydrothermal conditions. Astrobiology. 2008; 8(6): 1113-26
73. Mulkidjanian AY, Galperin MY. On the origin of life in the zinc world. 2. Validation of the hypothesis on the photosynthesizing zinc sulfide edifices as cradles of life on Earth. Biol Direct. 2009; 4: 27
74. Sleep NH. The Hadean-Archaean environment. Cold Spring Harb Perspect Biol. 2010; 2(6): a002527
75. Lisa M-N, Hemmingsen L, Vila AJ. Catalytic role of the metal ion in the metallo-{beta}-lactamase GOB. J Biol Chem. 2010; 28 (7): 4570-7
76. Li L, Lindquist S. Creating a protein-based element of inheritance. Science. 2000; 287(5453): 661-4
77. Uptain SM, Lindquist S. Prions as protein-based genetic elements. Annu Rev Microbiol. 2002; 56: 703-41
78. Koonin EV. Does the central dogma still stand? Biol Direct. 2012; 7: 27
79. Didonna A. Prion protein and its role in signal transduction. Cell Mol Biol Lett. 2013; 18(2): 209-30
80. Guerrier-Takada C, Gardiner K, Marsh T, Pace N, Altman S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell. 1983; 35(3 Pt 2): 849-57
81. Cech TR. RNA splicing: three themes with variations. Cell. 1983: 34(3): 713-6
82. Kurakin A. The self-organizing fractal theory as a universal discovery method: the phenomenon of life. Theor Biol Med Model 2011; 8: 4
83. Nobelprize.org. The 2013 Nobel Prize in Physiology or Medicine—Press Release. Available from: http://www.nobelprize.org/nobel_prizes/medicine/leureates/2013/pree.html (Accessed 12-9-2013)
84. Manassa RP, McCormick MI. Social learning and acquired recognition of a predator by a marine fish. Anim Cogn. 2012 Mar 28. [PubMed]
85. Mitchell MD, McCormick MI, Ferrari MC, Chivers DP. Coral reef fish rapidly learn to identify multiple unknown predators upon recruitment to the reef. PLoS One. 2011; 6(1): e15764
86. Suen G, Teiling C, Li L, Holt C, Abouheif E, Bornberg-Bauer E, et al. The genome sequence of the leaf-cutter ant Atta cephalotes reveals insights into its obligate symbiotic lifestyle. PLoS Genet 2011; 7(2): e1002007
87. Barke J, Seipke RF, Grüschow S, Heavens D, Drou N, Bibb MJ, et al. A mixed community of actinomycetes produce multiple antibiotics for the fungus farming ant Acromyrmex octospinosus. BMC Biol. 2010; 8: 109
88. Viana DS, Gordo I, Sucena E, Moita MA. Cognitive and motivational requirements for the emergence of cooperation in a rat social game. PLoS One. 2010; 5(1): e8483
89. Banerjee K, Chabris CF, Johnson VE, Lee JJ, Tsao F, et al. General intelligence in another primate: individual differences across cognitive task performance in a new world monkey (Saguinus oedipus). PLoS One. 2009; 4(6): e5883
90. Elgier AM, Jakovcevic A, Barrera G, Mustaca AE, Bentosela M. Communication between domestic dogs (Canis familiaris) and humans: dogs are good learners. Behav Processes. 2009; 81(3): 402-8
91. Hochner B, Shomrat T, Fiorito G. The octopus: a model for a comparative analysis of the evolution of learning and memory mechanisms. Biol Bull. 2006; 210(3): 308-17
92. Freeberg TM. Social transmission of courtship behavior and mating preferences in brown-headed cowbirds, Molothrus ater. Learn Behav. 2004; 32(1): 122-30
93. Thomson RL, Forsman JT, Monkkonen M. Positive interactions between migrant and resident birds: testing the heterospecific attraction hypothesis. Oecologia. 2003; 134(3): 431-8
94. Anderson JR, Sallaberry P, Barbier H. Use of experimenter-given cues during object-choice tasks by capuchin monkeys. Animal Behav.1995; 49(1): 201-8
95. Fiorito G, Scotto P. Observational learning in Octopus vulgaris. Science. 1992; 256(5056): 545-7
96. Lynch M. Colloquium Papers: The frailty of adaptive hypotheses for the origins of organismal complexity. Proc Natl Acad Sci USA. 2007; 104 (Suppl 1): 8597-604
97. Mustonen V, Lassig M. Fitness flux and ubiquity of adaptive evolution. Proc Natl Acad Sci USA. 2010; 107(9): 4248-53
98. Salvucci E. Selfishness, warfare, and economics; or integration, cooperation, and biology. Front Cell Infect Microbiol. 2012; 2: 54
99. Humpherys D, Eggan K, Akutsu H, Hochedlinger K, Rideout WM III, Biniszkiewicz D, et al. Epigenetic instability in ES cells and cloned mice. Science. 2001; 293(5527): 95-7
100. Feder JL, Roethele JB, Filchak K, Niedbalski J, Romero-Severson J. Evidence for inversion polymorphism related to sympatric host race formation in the apple maggot fly, Rhagoletis pomonella. Genetics. 2003; 163(3): 939-53
101. Sun YH, Chen SP, Wang YP, Hu W, Zhu ZY. Cytoplasmic impact on cross-genus cloned fish derived from transgenic common carp (Cyprinus carpio) nuclei and goldfish (Carassius auratus) enucleated eggs. Biol Reprod. 2005; 72(3): 510-5
102. Fishman L, Willis JH. A cytonuclear incompatibility causes anther sterility in Mimulus hybrids. Evolution. 2006; 60(7): 1372-81
103. Bowles EJ, Campbell KH, St John JC. Nuclear transfer: preservation of a nuclear genome at the expense of its associated mtDNA genome(s). Curr Top Dev Biol. 2007; 77: 251-90
104. Leppala J, Savolainen O. Nuclear-cytoplasmic interactions reduce male fertility in hybrids of Arabidopsis lyrata subspecies. Evolution. 2011; 65(10): 2959-72.
105. Cohen J, Alikani M. The biological basis for defining bi-parental or tri-parental origin of offspring from cytoplasmic and spindle transfer. Reprod Biomed Online. 2013; 26(6): 535-7
106. Tartakoff AM, Aylyarov I, Jaiswal P. Septin-containing barriers control the differential inheritance of cytoplasmic elements. Cell Rep. 2013; 3(1): 223-36
107. Tachibana M, Amato P, Sparman M, Woodward J, Sanchis DM, Ma H, et al. Towards germline gene therapy of inherited mitochondrial diseases. Nature. 2013; 493(7434): 627-31
108. Maurel M-C, Kanellopoulos-Langevin C. Heredity-venturing beyond genetics. Biol Reprod. 2008; 79(1): 2-8
109. Yin S-d. Six models of pregnancy-associated eukaryotic cell transmission among fetus, mother and infant. The Journal of Theoretical Fimpology. 2013; 1(2): e-20120609-1-2-3. Available from: www.fimpology.com
110. Yin S-d. The female may play more roles in biological heredity than the male does: Female-relayed integral cell and cytoplasmic inheritance. The Journal of Theoretical Fimpology. 2013; 1(3): e-20120717-1-3-6. Available from: http://www.fimpology.com
111. Lim CT, Bershadsky A, Sheetz MP. Mechanobiology. J R Soc Interface. 2010; 7(Suppl 3): S291-3
112. Makale M. Cellular mechanobiology and cancer metastasis. Birth Defects Res C Embryo Today. 2007; 81(4): 329-43
113. Backhed F, Ley RE, Sonnenburg JL, Peterson DA, Gordon JI. Host-bacterial mutualism in the human intestine. Science. 2005; 307(5717): 1915-20
114. Haburchak DR. Microbial inhabitants of humans: their ecology and role in health and disease. JAMA. 2005; 293(12): 1522-3
115. Paliy O, Kenche H, Abernathy F, Michail S. High-throughput quantitative analysis of the human intestinal microbiota with a phylogenetic microarray. Appl Environ Microbiol 2009; 75(11): 3572-9
116. Rice, Stanley A. Encyclopedia of Evolution. Pp. 276-278. Facts on File, Inc. New York. USA. 2007
117. Ernst Mayr. The growth of biological thought: diversity, evolution, and inheritance. Pp. 566-570. Belknap Press. USA. 1982
118. Michod RE, Nedelcu AM, Roze D. Cooperation and conflict in the evolution of individuality. IV. Conflict mediation and evolvability in Volvox carteri. Biosystems. 2003; 69(2-3): 95-114
119. Paenke I, Jin Y, Branke J. Balancing population- and individual-level adaptation in changing environments. Adaptive Behavior. 2009; 17(2): 153-74
120. Kirschner M, Gerhart J. Evolvability. Proc Natl Acad Sci USA. 1998; 95(15): 8420-7
121. Poole AM, Phillips MJ, Penny D. Prokaryote and eukaryote evolvability. Biosystems. 2003; 69(2-3): 163-85
122. Michod RE, Nedelcu AM, Roze D. Cooperation and conflict in the evolution of individuality. IV. Conflict mediation and evolvability in Volvox carteri. Biosystems. 2003; 69(2-3): 95-114
123. Jones AG, Arnold SJ, Burger R. The mutation matrix and the evolution of evolvability. Evolution. 2007; 61(4): 727-45
124. Snell-Rood EC. Selective processes in development: Implications for the costs and benefits of phenotypic plasticity. Integr Comp Biol. 2012; 52(1): 31-42
125. Brown RL. What evolvability really is. Br J Philos Sci. 2013; 0 (2013) axt014v1-axt014
126. Palmer ME, Moudgil A, Feldman MW. Long-term evolution is surprisingly predictable in lattice proteins. J R Soc Interface. 2013; 10(82): 20130026
127. Hendrix RW. Bacteriophage HK 97: assembly of the capsid and evolutionary connections. Advance Virus Research. 2005; 64:1-14
128. Suttle CA. Viruses in the sea. Nature. 2005; 437(7057): 356-61
129. Angly FE, Felts B, Breitbart M, Salamon P, Edwards RA, Carlson C, et al. The marine viromes of four oceanic regions. PLoS Biol. 2006; 4(11): e368
130. Clokie MRJ, Mann NH. Marine cyanophages and light. Environmental Microbiology. 2006; 8(12): 2074-82
131. Koonin EV. Darwinian evolution in the light of genomics. Nucleic Acids Res. 2009; 37(4): 1011-34
132. Whitman WB, Coleman DC, Wiebe WJ. Prokaryotes: the unseen majority. Proc Natl Acad Sci USA. 1998; 95(12): 6578-83
133. World Population. http://www.worldmeters.info/world-population/ (Accessed 12-22-2013).
Copyright © 2013- by Cory H. E. R. & C. Inc. All Rights Reserved.