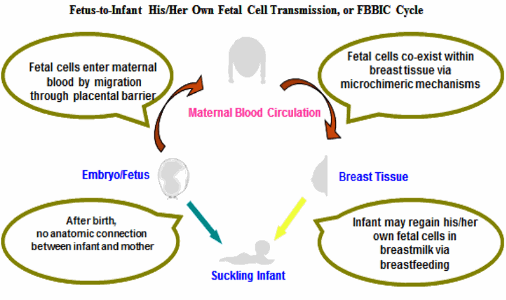
图: 妊娠期间,胎儿细胞在生理情况下突破胎盘屏障进入母体血液循环,并分布于包括乳腺在内的母体多个脏器和组织,部分胎儿细胞进入母乳中。尽管出生后,新生儿与母体之间不再存在任何解剖上的连接,如胎儿期的脐带和胎盘,母乳喂养可能扮演着一个替代角色。乳儿经由亲生母亲的母乳喂养,重新获得自己在胎儿期间储存在母乳中的胎儿细胞。
The Journal of Theoretical Fimpology. Volume 1, Issue 1: e-20120612-2. March 18, 2013 (www.fimpology.com)
ORCID ID: https://orcid.org/0009-0005-8661-6889
内容摘要
关键词: FBBIC 环路; 胎儿细胞; 胎儿; 婴儿; 母乳; 母乳喂养; 干细胞; 细胞迁移
导言
理论假说
图: 妊娠期间,胎儿细胞在生理情况下突破胎盘屏障进入母体血液循环,并分布于包括乳腺在内的母体多个脏器和组织,部分胎儿细胞进入母乳中。尽管出生后,新生儿与母体之间不再存在任何解剖上的连接,如胎儿期的脐带和胎盘,母乳喂养可能扮演着一个替代角色。乳儿经由亲生母亲的母乳喂养,重新获得自己在胎儿期间储存在母乳中的胎儿细胞。
支持性证据和讨论
I. 母体乳腺是胎儿细胞停留的组织和器官之一
II. 母乳中的细胞来自母体自身和胎儿
III. 母乳中的胎儿(崽)细胞穿过乳儿(崽)的胃肠道上皮组织并在乳儿(崽)体内重新分布
VI. FBBIC环路的可能意义和未来研究展望
妊娠期获得的细胞免疫经母乳喂养传递给新生乳儿
移植免疫学机制
干细胞对人乳喂养儿的再生保护作用
对母乳及母乳喂养的新认知
结束语
致谢
参考文献
1. Gammill HS, Nelson JL. Naturally acquired microchimerism. Int J Dev Biol. 2010; 54: 531–43
2. Bianchi DW, Zickwolf GK, Weil GJ, Sylvester S, DeMaria MA. Male fetal progenitor cells persist in maternal blood for as long as 27 years postpartum. Proc Natl Acad Sci USA. 1996; 93: 705–8
3. Khosrotehrani K, Johnson KL, Cha DH, Salomon RN, Bianchi DW. Transfer of fetal cells with multilineage potential to maternal tissue. JAMA. 2004; 292: 75–80
4. Vernochet C, Caucheteux SM, Kanellopoulos-Langevin C. Bi-directional cell trafficking between mother and fetus in mouse placenta. Placenta. 2007; 28(7): 639–49
5. Stevens AM, Hermes HM, Kiefer MM, Rutledge JC, Nelson JL. Chimeric maternal cells with tissue-specific antigen expression and morphology are common in infant tissues. Pediatr Dev Pathol. 2009; 12(5): 337–46
6. Karniychuk UU, Van Breedam W, Van Roy N, Rogel-Gaillard C, Nauwynck HJ. Demonstration of microchimerism in pregnant sows and effects of congenital PRRSV infection. Vet Res. 2012; 43(1): 19
7. McConico A, Butters K, Lien K, Knudsen B, Wu X, Platt JL, et al. In utero cell transfer between porcine littermates. Reprod Fertil Dev. 2011; 23(2): 297–302
8. Lambert NC, Pang JM, Yan Z, Erickson TD, Stevens AM, Furst DE, et al. Male microchimerism in women with systemic sclerosis and healthy women who have never given birth to a son. Ann Rheum Dis. 2005; 64(6): 845–8
9. Yan Z, Lambert NC, Guthrie KA, Porter AJ, Loubiere LS, Madeleine MM, et al. Male microchimerism in women without sons: quantitative assessment and correlation with pregnancy history. Am J Med. 2005; 118(8): 899–906
10. van Dijk BA, Boomsma DI, de Man AJ. Blood group chimerism in human multiple births is not rare. Am J Med Genet. 1996; 61: 264–8
11. Ma LJ, Walter B, Deguzman A, Muller HK, Walker AM. Trans-epithelial immune cell transfer during suckling modulates delayed-type hypersensitivity in recipients as a function of gender. PLoS One. 2008; 3: e3562
12. Zhou L, Yoshimura Y, Huang Y, Suzuki R, Yokoyama M, Okabe M, et al. Two independent pathways of maternal cell transmission to offspring: through placenta during pregnancy and by breast-feeding after birth. Immunology. 2000; 101: 570–80
13. Williams PP. Immunomodulating effects of intestinal absorbed maternal colostral leukocytes by neonatal pigs. Can J Vet Res. 1993; 57: 1–8
14. Tuboly S, Bernath S, Glavits R, Medveczky I. Intestinal absorption of colostral lymphoid cells in newborn piglets. Vet Immunol Immunopathol. 1988; 20: 75–85
15. Sunami R, Komuro M, Tagaya H, Hirata S. Migration of microchimeric fetal cells into maternal circulation before placenta formation. Chimerism. 2010; 1: 66–8
16. Fujiki Y, Johnson KL, Peter I, Tighiouart H, Bianchi DW. Fetal cells in the pregnant mouse are diverse and express a variety of progenitor and differentiated cell markers. Biol Reprod. 2009; 81: 26–32
17. O’Donoghue K, Chan J, de la Fuente J, Kennea N, Sandison A, Anderson JR, et al. Microchimerism in female bone marrow and bone decades after fetal mesenchymal stem-cell trafficking in pregnancy. Lancet. 2004; 364:179–82
18. Wegmann TG, Lin H, Guilbert L, Mosmann TR. Bidirectional cytokine interactions in the maternal-fetal relationship: is successful pregnancy a TH2 phenomenon? Immunology Today. 1993; 14: 353–6
19. Jeurink PV, van Bergenhenegouwen J, Jimenez E, Knippels LM, Fernandez L, Garssen J, et al. Human milk: a source of more life than we imagine. Benef Microbes. Epub 2012 Dec 27:17-30. PubMed PMID: 23271066
20. Cabrera-Rubio R, Collado MC, Laitinen K, Salminen S, Isolauri E, Mira A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am J Clin Nutr. 2012; 96(3): 544–51
21. Hunt KM, Foster JA, Forney LJ, Schutte UM, Beck DL, Abdo Z, et al. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS One. 2011; 6(6): e21313
22. Collado MC, Delgado S, Maldonado A, Rodríguez JM. Assessment of the bacterial diversity of breast milk of healthy women by quantitative real-time PCR. Lett Appl Microbiol. 2009; 48(5): 523–8
23. Martín R, Heilig HG, Zoetendal EG, Jimenez E, Fernandez L, Smidt H, et al. Cultivation-independent assessment of the bacterial diversity of breast milk among healthy women. Res Microbiol. 2007; 158(1): 31–7
24. Perez PF, Dore J, Leclerc M, Levenez F, Benyacoub J, Serrant P, et al. Bacterial imprinting of the neonatal immune system: lessons from maternal cells? Pediatrics. 2007; 119(3): e724–32
25. Smith CW, Goldman AS. The cells of human colostrum. I. In vitro studies of morphology and functions. Pediatr Res. 1968; 2: 103–9
26. Wirt DP, Adkins LT, Palkowetz KH, Schmalstieg FC, Goldman AS. Activated and memory T lymphocytes in human milk. Cytometry. 1992; 13: 282–90
27. Tuaillon E, Valea D, Becquart P, Al Tabaa Y, Meda N, Bollore K, et al. Human milk-derived B cells: a highly activated switched memory cell population primed to secrete antibodies. J Immunol. 2009; 182: 7155–62
28. Jain L, Vidyasagar D, Xanthou M, Ghai V, Shimada S, Blend M. In vivo distribution of human milk leucocytes after ingestion by newborn baboons. Arch Dis Child. 1989; 64: 930–3
29. Fan Y, Chong YS, Choolani MA, Cregan MD, Chan JK. Unravelling the mystery of stem/progenitor cells in human breast milk. PLoS One. 2010; 5: e14421
30. Crago SS, Prince SJ, Pretlow TG, McGhee JR, Mestecky J. Human colostral cells. I. Separation and characterization. Clin Exp Immunol. 1979; 38: 585–97
31. Cregan MD, Fan Y, Appelbee A, Brown ML, Klopcic B, Koppen J, et al. Identification of nestin-positive putative mammary stem cells in human breastmilk. Cell Tissue Res. 2007; 329: 129–136
32. Patki S, Kadam S, Chandra V, Bhonde R. Human breast milk is a rich source of multipotent mesenchymal stem cells. Hum Cell. 2010; 23(2): 35–40
33. Indumathi S, Dhanasekaran M, Rajkumar JS, Sudarsanam D. Exploring the stem cell and non-stem cell constituents of human breast milk. Cytotechnology. Epub 2012 Sep 2. PubMed PMID: 22940915
34. Shevde N. Stem cells: flexible friends. Nature. 2012; 483: S22–6
35. Weiler IJ, Hickler W, Sprenger R. Demonstration that milk cells invade the suckling neonatal mouse. Am J Reprod Immunol. 1983; 4: 95–8
36. Geddes L. Breast-milk stem cells may bypass ethical dilemmas. New Scientist. 2011 November 16. Available from: http://www.newscientist.com/article/mg21228392.600-breastmilk-stem-cells-may-bypass-ethical-dilemmas...
37. Schlesinger JJ, Covelli HD. Evidence for transmission of lymphocyte responses to tuberculin by breast feeding. The Lancet. 1977; 310(8037): 529–32
38. Bandrick M, Pieters M, Pijoan C, Molitor TW. Passive transfer of maternal Mycoplasma hyopneumoniae-specific cellular immunity to piglets. Clin Vaccine Immunol. 2008; 15(3): 540–43
39. Sabbaj S, Ghosh MK, Edwards BH, Leeth R, Decker WD, Goepfert PA, et al. Breast milk-derived antigen-specific CD8+ T cells: an extralymphoid effector memory cell population in humans. J Immunol. 2005; 174(5): 2951–6
40. Permar SR, Kang HH, Carville A, Wilks AB, Mansfield KG, Rao SS, et al. Preservation of memory CD4(+) T lymphocytes in breast milk of lactating rhesus monkeys during acute simian immunodeficiency virus infection. J Infect Dis. 2010; 201(2): 302–10
41. Andrassy J, Kusaka S, Jankowska-Gan E, Torrealba JR, Haynes LD, Marthaler BR, et al. Tolerance to noninherited maternal MHC antigens in mice. J Immunol. 2003; 171(10): 5554–61
42. Muller YD, Seebach JD, Bühler LH, Pascual M, Golshayan D. Transplantation tolerance: Clinical potential of regulatory T cells. Self Nonself. 2011; 2(1): 26–34
43. Mold JE, Michaelsson J, Burt TD, Muench MO, Beckerman KP, Busch MP, et al. Maternal alloantigens promote the development of tolerogenic fetal regulatory T cells in utero. Science. 2008; 322: 1562–5
44. Dutta P, Molitor-Dart M, Bobadilla JL, Roenneburg DA, Yan Z, Torrealba JR, et al. Microchimerism is strongly correlated with tolerance to noninherited maternal antigens in mice. Blood. 2009; 114: 3578–87
45. Hasselbalch H, Jeppesen DL, Engelmann MD, Michaelsen KF, Nielsen MB. Decreased thymus size in formula-fed infants compared with breastfed infants. Acta Paediatr. 1996; 85: 1029–32
46. Jeppesen D, Hasselbalch H, Ersboll AK, Heilmann C, Valerius NH. Thymic size in uninfected infants born to HIV-positive mothers and fed with pasteurized human milk. Acta Paediatr. 2003; 92: 679–83
47. Jeppesen DL, Hasselbalch H, Lisse IM, Ersboll AK, Engelmann MD. T-lymphocyte subsets, thymic size and breastfeeding in infancy. Pediatr Allergy Immunol. 2004; 15: 127–32
48. Santos MA, O'Donoghue K, Wyatt-Ashmead J, Fisk NM. Fetal cells in the maternal appendix: a marker of inflammation or fetal tissue repair? Hum Reprod. 2008; 23: 2319–25
49. Mikhail MA, M'Hamdi H, Welsh J, Levicar N, Marley SB, Nicholls JP, et al. High frequency of fetal cells within a primitive stem cell population in maternal blood. Hum Reprod. 2008; 23(4): 928–33
50. O'Donoghue K, Sultan HA, Al-Allaf FA, Anderson JR, Wyatt-Ashmead J, Fisk NM. Microchimeric fetal cells cluster at sites of tissue injury in lung decades after pregnancy. Reprod Biomed Online. 2008;16: 382–90
51. Rossi G. Nature of stem cells involved in fetomaternal microchimerism. Lancet. 2004; 364: 1936
52. Khosrotehrani K, Johnson KL, Cha DH, Salomon RN, Bianchi DW. Transfer of fetal cells with multilineage potential to maternal tissue. JAMA. 2004; 292: 75–80
53. Srivatsa B, Srivatsa S, Johnson KL, Samura O, Lee SL, Bianchi DW. Microchimerism of presumed fetal origin in thyroid specimens from women: a case-control study. Lancet. 2001; 358: 2034–8
54. Oddy WH, Sly PD, de Klerk NH, Landau LI, Kendall GE, Holt PG, et al. Breast feeding and respiratory morbidity in infancy: a birth cohort study. Arch Dis Child. 2003; 88: 224–8
55. Chantry CJ, Howard CR, Auinger P. Full breastfeeding duration and associated decrease in respiratory tract infection in US children. Pediatrics. 2006; 117: 425–32
56. Duijts L, Jaddoe VWV, Hofman A, Moll HA. Prolonged and exclusive breastfeeding reduces the risk of infectious diseases in infancy. Pediatrics. 2010; 126:1 e18–e25
57. Chisti MJ, Salam MA, Smith JH, Ahmed T, Ashraf H, Bardhan PK, et al. Impact of lack of breast feeding during neonatal age on the development of clinical signs of pneumonia and hypoxemia in young infants with diarrhea. PLoS One. 2011; 6: e25817
58. Hoddinott P, Tappin D, Wright C. Breast feeding. BMJ. 2008; 336(7649): 881–7
59. Section on breastfeeding. Breastfeeding and the use of human milk. Pediatrics. 2012; 129(3): e827–41