The existing literature has recently indicated that (i) fetal cells are present in maternal
blood and other multiple maternal tissues and organs during and after pregnancy; (ii)
different functional immune cells and stem cells have been found in human breastmilk;
and (iii) intact animal milk cells can migrate into the gut wall of suckling offspring and
distribute further over the animal neonatal body. Based on these advantages, I propose a
novel model called the “Fetus-Breastmilk-Breastfeeding-Infant-Cells Cycle” (FBBIC
Cycle), or “the External Transmission of His/Her Own Fetal Cell from Fetus to Infant,”
to reveal an unrecognized mechanism in which some stem cells and functional immune
cells derived from embryonic/fetal cells become part of milk cells and can be ingested by
breastfed infants; these regained fetal cells can then be redistributed within the suckling
body without potential allogenic or semi-allogenic problems. The completion of the
FBBIC Cycle depends on the continuity from gestation to lactation. The FBBIC Cycle
theory suggested for the first time that the fetus may contribute milk cells and that
acquired cellular immunity during pregnancy may be transferred to the suckling via the
FBBIC Cycle. The potential benefits of the FBBIC Cycle in immunology and
regenerative cytology warrant further investigation.
It is well known that fetal growth and development within the uterus absolutely
depends on nurturing and anti-infection protection supplied by the maternal body through
diverse maternal-fetal molecule transferring mechanisms. Traditionally, it has been
assumed that the placental barrier might structurally and functionally isolate the
eukaryotic cells of the maternal body from those of the fetus [1] and that the cellular
component in breast milk was believed to be derived only from maternal host cell
lineages, in which fetal cells did not participate.
During the past two decades, accumulating evidence from studies on humans and
non-human mammals has indicated that the efficiency of the placental barrier for the
structural and functional isolation between maternal cells and fetal cells is relative and
that there is a physiologically bidirectional cell transmission between pregnant mothers
and their fetuses.[1-5] Moreover, the cell transmission between fetuses, also known as
"transmaternal sibling microchimerism,"[6-10] and the cellular migration between the
lactating mother and suckling offspring [11-14] have also been proposed.
Hypothesis
In this article, a novel model called the "Fetus-Breastmilk-Breastfeeding-Infant-Cells
Cycle" (FBBIC Cycle), or "the External Transmission of His/Her Own Fetal Cell from
Fetus to Infant," is hypothesized (see Figure), which consists of the following three major
statements: (i) Maternal breasts should be one of the organs where fetal cells can reside.
Fetal cells can colonize maternal mammary glands after migration through the placental
barrier and into maternal blood; (ii) Milk cells consist of heterogeneous cell lineages
from both maternal and fetal sources. Fetal cells in breast milk are expected to consist of
stem/progenitor cells and diverse end-differentiated cells including leukocytes; and (iii)
Regained fetal cells may reallocate within the suckling body. Fetal cells in breast milk
have the capability to cross the intestinal epithelium. After crossing the epithelium, fetal
cells may re-spread further and take up residence within a variety of organs and tissues of
the suckling without allogenic and semi-allogenic problems.
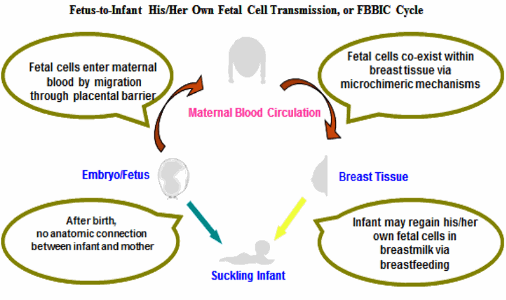
Figure:Figure. During gestation, fetal cells enter maternal blood by migrating through the placental barrier, and then spread into multiple maternal tissues and organs including breast tissue and breast milk, where fetal cells co-exist via microchimeric mechanisms. Although the prenatal anatomic connection (umbilical cord and placenta) no longer exists between the infant and mother during lactation, breastfeeding may replace its role. The suckling infant may be able to regain his/her fetal cells in breast milk via breastfeeding from his/her own lactating mother.
Supporting Evidence and Discussion
I. Maternal mammary glands as one of residing organs for fetal cells
Fetal cells, including end-differentiated cells and stem/progenitor cells, are normally
present in maternal bloodstream and many other maternal tissues and organs during
pregnancy and after delivery;[1-3, 15, 16] and, moreover, fetal cells can persist in the
maternal body for as long as 27 years, [2] or even for life.[17]
Interestingly, to the best of my knowledge, the question of whether fetal cells colonize
maternal mammary glands has not yet been answered. Moreover, the newly understood
fact that normal pregnancy is characterized by a maternal systemic Th2 immune response
[18] and that bacterial microorganisms exist in normal breast milk,[19-24] leads to the
hypothesis that maternal breasts may be one of the organs where fetal cells can be recruited to and reside.
II. Milkcells consisting of heterogeneous cell lineages from both maternal and fetal sources
Milkcells have been assumed for a long time to come only from diverse maternal end-
differentiated cell lineages, including phagocytes,[25] immunologic memory T cells [26]
and B cells.[27] The concentration of milkcells varies widely from 103 to 108 cells per mL
of human milk. [28-30] Recently, human milk cells bearing stem cell markers have been
identified by several research teams.[29, 31-33] Embryonic stem cells are defined as cells
that come from an organism at its earliest stages of development,[34] and can infinitely
renew themselves and transform into multicellular lineages in the body of
macroorganisms such as human beings and non-human mammals.[3, 34] However, the
source of stem cells in breast milk is elusive, although it was assumed to be from maternal mammary stem cells, not from the fetus.
In the FBBIC Cycle theory, it is hypothesized that (i) milk cells may be derived not
only from maternal host cells but also from fetal cells; (ii) the concentration of fetal cells
in breast milk is expected to reach its highest level in colostrum and then decline after
delivery; and (iii) fetal cells in breast milk may consist of haemopoietic or/and
mesenchymal stem cells and diverse end-differentiated cells, including leukocytes.
III. Re-spreading of fetal cells within the body of suckling offspring via breastfeeding
There has been a notion since the 1980s that intact leukocytes in ingested maternal
colostrum and mature milk can migrate into the gut wall of suckling offspring. Although,
up to now, no direct evidence from studies on human beings is available, many relevant
data from studies on non-human mammals have revealed that the trans-epithelial milk
cells truly exist within the body of animal suckling neonate.[11-14]
The concentration of milkcells in human colostrum is reported to be as high as 108 per
ml of milk.[28] Given that human neonates ingest on average 100 milliliters of colostrum
per day during the first 3 postnatal days, the total daily acquired cells from maternal
breastfeeding would be estimated to be as high as 1010 cells. If the trans-epithelium rate
of ingested milk cells in human neonates is 0.1%,[35] the total colostral cells regained by a
neonatal body would reach 107 cells per day. If two per cent of milk cells were stem
cells,[36] neonates would regain around 105 stem cells from colostrum-feeding daily.
Tuboly and colleagues described a phenomenon in the 1980s that the digestive tract
mucosal barrier of suckling newborn piglets was selectively opened to the colostral
lymphoid cells from their biological mother, but not to the cells from allogeneic lactating
mother, or from blood and heat-treated colostral lymphoid cells.[14] I argue that the
Tuboly Phenomenon may also occur in humans.
The intestinal epithelium-transferred animal milk cells have been found to redistribute
over the thymus, liver, lung, lymph nodes, spleen and gastrointestinal tissues of the
animal suckling.[11-14] Traditionally, these milk cells within the body of animal offspring
were also assumed to come only from the animal maternal host cells.
In the FBBIC Cycle theory, milk cells are hypothesized to come from both of maternal
and fetal sources, and therefore, it could be deducted that epithelium-transferred and then
body-redistributed milk cells may include animal embryonic/fetal cells, from which the
third hypothesis may be further inferred: Embryonic/fetal cells in human milk may re-
spread within the body of suckling infant via breastfeeding.
VI. Potential roles and significances of FBBIC Cycle and future prospects
The following is a brief introduction to the potential roles and significances of the FBBIC Cycle in the transmission of cellular immunity to suckling neonates, the
immunological mechanisms of transplantation, the regenerative effect of stem cells in human breastfed offspring, and our understanding of human milk and breastfeeding.
Transmission of cellular immunity to suckling neonates
Some studies have already suggested the possible roles played by the FBBIC Cycle in
the transmission of cellular immunity to suckling neonates. For instance, three decades
ago, breastfeeding was found to deliver a T lymphocyte-mediated immune response to
the BCG vaccine in breastfed infants.[37] Recently, vaccine-induced Mycoplasma
hyopneumoniae cellular immunity at 5 and 3 weeks antepartum was shown to be
transferred from vaccinated sows to their offspring via breastfeeding.[38] The transferred
immune cells were assumed to be of animal maternal origin.[37, 38]
However, according to the FBBIC Cycle theory, during gestation, fetal cells can
migrate into the maternal blood circulation and multiple other tissues or organs where fetal cells have an opportunity to be exposed to novel antigens that the pregnant mother may be experiencing. Some fetal cells have embryonic stem cell-like functions and can differentiate into different cell lineages, including immune cells (such as memory T cells).
If these fetal cells return to the body of a breastfed infant, the acquired cellular immunity would undoubtedly be transferred to the suckling. Therefore, the potential role of human fetal cells in generating tuberculin-specific cellular immunity and the specific cellular immunity for Mycoplasma hyopneumoniae obtained by animal fetal cells during gestation
cannot be excluded.
Recently, CD8(+) or CD4(+) T cells for virus-specific cellular immune responses
have been detected in both human and animal milk samples.[39, 40] The question of
whether the FBBIC Cycle plays a role in the transmission of virus-specific cellular
immunity to suckling neonates needs to be answered in the future.
Transplantation immunology
The survival of grafts at the individual macroorganism level involves a complicated
tolerogenic mechanism, one aspect of which is the tolerance to noninherited maternal
antigens (NIMA) mediated by maternal-fetal-microchimerism. [41] Recently, the role of
human regulatory T cells (Tregs) in transplantation tolerance has been attracting much
attention.[42] The development of tolerogenic fetal Tregs during gestation was found to be
enhanced by maternal alloantigens, and these tolerogenic fetal Tregs play a role in the
mechanism of tolerance to NIMA. [43, 44] In addition, many other cell lineages, such as
mesenchymal stem cells, embryonic stem cells, myeloid-derived suppressor cells,
tolerogenic DC, and B cells, have also been shown to have immuno-regulatory
properties.[42]
Based on Mold and colleagues' finding,[43] the fetus-to-fetus eukaryotic cell
transmission model and the FBBIC Cycle, the following possibility cannot be excluded:
Paternal alloantigens-tolerogenic fetal T regulatory cells (PATF-TRCs) may also be
induced during gestation, in addition to maternal alloantigens-tolerogenic fetal T
regulatory cells (MATF-TRCs). Once these MATF-TRCs and PATF-TRCs return to the
body of a suckling infant via breastfeeding, they survive much more easily than allogenic
and semi-allogenic cells do. These fetal cell-derived MATF-TRCs and PATF-TRCs may
participate in suppressing postnatal anti-maternal or anti-paternal immune reactions to
maternal or paternal grafting.
Regenerative cytological role in human breastfed offspring
The ultrasound assessment of the thymus revealed that the thymus size of breastfed
infants is significantly larger than that of formula-fed infants.[45-47] Jeppesen and
colleagues showed that thymus index and thymic index/weight-ratio of healthy infants
who were fed exclusively with pasteurized donor milk were significantly lower than
those of exclusively breastfed infants,[46] which cannot be easily explained by the
difference in the molecular components between the two kinds of human breastmilk.
However, it can be accounted for by the lack of the FBBIC Cycle in those infants fed
with pasteurized donor milk.
Studies on human beings and nonhuman animal models have shown that fetal cells
may be involved in maternal tissue repair.[48-53] However, according to the FBBIC Cycle
theory, the critical question of whether regained fetal cells participate in the tissue repair
of breastfed infants also has clinical significance. The potential regenerative protection
from regained fetal cells through the FBBIC Cycle may provide a novel mechanism for
explaining the finding in epidemiological studies that breastfeeding decreases the infant
mortality and morbidity from infections in the gastrointestinal tract or respiratory tract.[54-
57]
Significance in our understanding on human milk and breastfeeding
To date, our understanding on human milk and breastfeeding has focused on the
aspects of nutrition and immunology: (i) at the nutritional level, breastmilk offers various
essential nutrients including carbohydrates, lipids and proteins, for the growth and
development of suckling offspring; and (ii) at the immunological level, breastmilk acts as
a vector to transfer maternal acquired and innate immune defense factors to suckling
babies. The FBBIC cycle may further suggest that the fetal cellular benefit of human milk
and breastfeeding for infants to obtain from their healthy biological mothers is
individually unique. It is may be true that discarding biological mother's breastfeeding
means abandoning the chance for offspring to regain their own fetal stem/progenitor cells
and functional immune cells that were stored in breastmilk during pregnancy.
Breastfeeding has been consistently advocated as the best feeding method for human
suckling by academic institutes and international organizations, such as WHO and
UNICEF, over the past decades.[58; 59] The FBBIC Cycle may provide a new theoretical
basis for supporting human breastfeeding, especially colostral feeding.
Conclusion
During the past decades, the advantages in cellular embryology, fetology, regenerative
cytology, immunology, and human milk research have presented many indirect pieces of
evidence supporting the FBBIC Cycle theory. For the first time, the FBBIC Cycle theory
explains theoretically that the fetus may play an active role, or be a co-contributor, in the
formation of maternal breast milk, and breastfed infants may regain their own fetal cells
from ingested breastmilk.
The FBBIC Cycle is one of the pregnancy-induced cell transmissions that occur
among the fetus, infant, and the maternal body, and its completion absolutely depends on
the continuity from gestation to lactation, meaning that the FBBIC Cycle may not be
completed in infants who are never breastfed by their own mothers. The probably
irreplaceable benefits of the FBBIC Cycle for infants in immunology and regenerative
cytology are worth further investigation, which will undoubtedly improve our
understanding of human breast milk and breastfeeding.
Acknowledgments
I express my thanks to Professor Qian Qian, Professor Ronghua Chen, Professor
Shiyuan Chen and Professor Xinyou Jiang of The First Affiliated Hospital and The
Second Affiliated Hospital of Nanjing Medical University, as well as Professor Walker
and Dr. Israel of Harvard Medical School and Massachusetts General Hospital, and
Professor Simister of Brandeis University for their giving me the opportunity in the
domain of human milk research. I thank Professor Nelson, Dr. Burlingham and two
anonymous reviewers for their critical reading and comments on the previous versions between June and November, 2012. Also, I am grateful to my father, mother Yulan Ju,
wife and children for their support and patience. I have to thank Mr. W. Liu and Mr. J.
Dong for their technical help, and many staff at the University of British Columbia
Library, Simon Fraser University Library, Vancouver Public Library, and Burnaby Public
Library for their assistance in my search for references.
Reference
1. Gammill HS, Nelson JL. Naturally acquired microchimerism. Int J Dev Biol. 2010; 54: 531–43
2. Bianchi DW, Zickwolf GK, Weil GJ, Sylvester S, DeMaria MA. Male fetal progenitor cells persist in maternal blood for as long as 27 years postpartum. Proc Natl Acad Sci USA. 1996; 93: 705–8
3. Khosrotehrani K, Johnson KL, Cha DH, Salomon RN, Bianchi DW. Transfer of fetal cells with multilineage potential to maternal tissue. JAMA. 2004; 292: 75–80
4. Vernochet C, Caucheteux SM, Kanellopoulos-Langevin C. Bi-directional cell trafficking between mother and fetus in mouse placenta. Placenta. 2007; 28(7): 639–49
5. Stevens AM, Hermes HM, Kiefer MM, Rutledge JC, Nelson JL. Chimeric maternal cells with tissue-specific antigen expression and morphology are common in infant tissues. Pediatr Dev Pathol. 2009; 12(5): 337–46
6. Karniychuk UU, Van Breedam W, Van Roy N, Rogel-Gaillard C, Nauwynck HJ. Demonstration of microchimerism in pregnant sows and effects of congenital PRRSV infection. Vet Res. 2012; 43(1): 19
7. McConico A, Butters K, Lien K, Knudsen B, Wu X, Platt JL, et al. In utero cell transfer between porcine littermates. Reprod Fertil Dev. 2011; 23(2): 297–302
8. Lambert NC, Pang JM, Yan Z, Erickson TD, Stevens AM, Furst DE, et al. Male microchimerism in women with systemic sclerosis and healthy women who have never given birth to a son. Ann Rheum Dis. 2005; 64(6): 845–8
9. Yan Z, Lambert NC, Guthrie KA, Porter AJ, Loubiere LS, Madeleine MM, et al. Male microchimerism in women without sons: quantitative assessment and correlation with pregnancy history. Am J Med. 2005; 118(8): 899–906
10. van Dijk BA, Boomsma DI, de Man AJ. Blood group chimerism in human multiple births is not rare. Am J Med Genet. 1996; 61: 264–8
11. Ma LJ, Walter B, Deguzman A, Muller HK, Walker AM. Trans-epithelial immune cell transfer during suckling modulates delayed-type hypersensitivity in recipients as a function of gender. PLoS One. 2008; 3: e3562
12. Zhou L, Yoshimura Y, Huang Y, Suzuki R, Yokoyama M, Okabe M, et al. Two independent pathways of maternal cell transmission to offspring: through placenta during pregnancy and by breast-feeding after birth. Immunology. 2000; 101: 570–80
13. Williams PP. Immunomodulating effects of intestinal absorbed maternal colostral leukocytes by neonatal pigs. Can J Vet Res. 1993; 57: 1–8
14. Tuboly S, Bernath S, Glavits R, Medveczky I. Intestinal absorption of colostral lymphoid cells in newborn piglets. Vet Immunol Immunopathol. 1988; 20: 75–85
15. Sunami R, Komuro M, Tagaya H, Hirata S. Migration of microchimeric fetal cells into maternal circulation before placenta formation. Chimerism. 2010; 1: 66–8
16. Fujiki Y, Johnson KL, Peter I, Tighiouart H, Bianchi DW. Fetal cells in the pregnant mouse are diverse and express a variety of progenitor and differentiated cell markers. Biol Reprod. 2009; 81: 26–32
17. O’Donoghue K, Chan J, de la Fuente J, Kennea N, Sandison A, Anderson JR, et al. Microchimerism in female bone marrow and bone decades after fetal mesenchymal stem-cell trafficking in pregnancy. Lancet. 2004; 364:179–82
18. Wegmann TG, Lin H, Guilbert L, Mosmann TR. Bidirectional cytokine interactions in the maternal-fetal relationship: is successful pregnancy a TH2 phenomenon? Immunology Today. 1993; 14: 353–6
19. Jeurink PV, van Bergenhenegouwen J, Jimenez E, Knippels LM, Fernandez L, Garssen J, et al. Human milk: a source of more life than we imagine. Benef Microbes. Epub 2012 Dec 27:17-30. PubMed PMID: 23271066
20. Cabrera-Rubio R, Collado MC, Laitinen K, Salminen S, Isolauri E, Mira A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am J Clin Nutr. 2012; 96(3): 544–51
21. Hunt KM, Foster JA, Forney LJ, Schutte UM, Beck DL, Abdo Z, et al. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS One. 2011; 6(6): e21313
22. Collado MC, Delgado S, Maldonado A, Rodríguez JM. Assessment of the bacterial diversity of breast milk of healthy women by quantitative real-time PCR. Lett Appl Microbiol. 2009; 48(5): 523–8
23. Martín R, Heilig HG, Zoetendal EG, Jimenez E, Fernandez L, Smidt H, et al. Cultivation-independent assessment of the bacterial diversity of breast milk among healthy women. Res Microbiol. 2007; 158(1): 31–7
24. Perez PF, Dore J, Leclerc M, Levenez F, Benyacoub J, Serrant P, et al. Bacterial imprinting of the neonatal immune system: lessons from maternal cells? Pediatrics. 2007; 119(3): e724–32
25. Smith CW, Goldman AS. The cells of human colostrum. I. In vitro studies of morphology and functions. Pediatr Res. 1968; 2: 103–9
26. Wirt DP, Adkins LT, Palkowetz KH, Schmalstieg FC, Goldman AS. Activated and memory T lymphocytes in human milk. Cytometry. 1992; 13: 282–90
27. Tuaillon E, Valea D, Becquart P, Al Tabaa Y, Meda N, Bollore K, et al. Human milk-derived B cells: a highly activated switched memory cell population primed to secrete antibodies. J Immunol. 2009; 182: 7155–62
28. Jain L, Vidyasagar D, Xanthou M, Ghai V, Shimada S, Blend M. In vivo distribution of human milk leucocytes after ingestion by newborn baboons. Arch Dis Child. 1989; 64: 930–3
29. Fan Y, Chong YS, Choolani MA, Cregan MD, Chan JK. Unravelling the mystery of stem/progenitor cells in human breast milk. PLoS One. 2010; 5: e14421
30. Crago SS, Prince SJ, Pretlow TG, McGhee JR, Mestecky J. Human colostral cells. I. Separation and characterization. Clin Exp Immunol. 1979; 38: 585–97
31. Cregan MD, Fan Y, Appelbee A, Brown ML, Klopcic B, Koppen J, et al. Identification of nestin-positive putative mammary stem cells in human breastmilk. Cell Tissue Res. 2007; 329: 129–136
32. Patki S, Kadam S, Chandra V, Bhonde R. Human breast milk is a rich source of multipotent mesenchymal stem cells. Hum Cell. 2010; 23(2): 35–40
33. Indumathi S, Dhanasekaran M, Rajkumar JS, Sudarsanam D. Exploring the stem cell and non-stem cell constituents of human breast milk. Cytotechnology. Epub 2012 Sep 2. PubMed PMID: 22940915
35. Weiler IJ, Hickler W, Sprenger R. Demonstration that milk cells invade the suckling neonatal mouse. Am J Reprod Immunol. 1983; 4: 95–8
36. Geddes L. Breast-milk stem cells may bypass ethical dilemmas. New Scientist. 2011 November 16. Available from: http://www.newscientist.com/article/mg21228392.600-breastmilk-stem-cells-may-bypass-ethical-dilemmas...
37. Schlesinger JJ, Covelli HD. Evidence for transmission of lymphocyte responses to tuberculin by breast feeding. The Lancet. 1977; 310(8037): 529–32
38. Bandrick M, Pieters M, Pijoan C, Molitor TW. Passive transfer of maternal Mycoplasma hyopneumoniae-specific cellular immunity to piglets. Clin Vaccine Immunol. 2008; 15(3): 540–43
39. Sabbaj S, Ghosh MK, Edwards BH, Leeth R, Decker WD, Goepfert PA, et al. Breast milk-derived antigen-specific CD8+ T cells: an extralymphoid effector memory cell population in humans. J Immunol. 2005; 174(5): 2951–6
40. Permar SR, Kang HH, Carville A, Wilks AB, Mansfield KG, Rao SS, et al. Preservation of memory CD4(+) T lymphocytes in breast milk of lactating rhesus monkeys during acute simian immunodeficiency virus infection. J Infect Dis. 2010; 201(2): 302–10
41. Andrassy J, Kusaka S, Jankowska-Gan E, Torrealba JR, Haynes LD, Marthaler BR, et al. Tolerance to noninherited maternal MHC antigens in mice. J Immunol. 2003; 171(10): 5554–61
42. Muller YD, Seebach JD, Bühler LH, Pascual M, Golshayan D. Transplantation tolerance: Clinical potential of regulatory T cells. Self Nonself. 2011; 2(1): 26–34
43. Mold JE, Michaelsson J, Burt TD, Muench MO, Beckerman KP, Busch MP, et al. Maternal alloantigens promote the development of tolerogenic fetal regulatory T cells in utero. Science. 2008; 322: 1562–5
44. Dutta P, Molitor-Dart M, Bobadilla JL, Roenneburg DA, Yan Z, Torrealba JR, et al. Microchimerism is strongly correlated with tolerance to noninherited maternal antigens in mice. Blood. 2009; 114: 3578–87
46. Jeppesen D, Hasselbalch H, Ersboll AK, Heilmann C, Valerius NH. Thymic size in uninfected infants born to HIV-positive mothers and fed with pasteurized human milk. Acta Paediatr. 2003; 92: 679–83
48. Santos MA, O'Donoghue K, Wyatt-Ashmead J, Fisk NM. Fetal cells in the maternal appendix: a marker of inflammation or fetal tissue repair? Hum Reprod 2008; 23: 2319–25
49. Mikhail MA, M'Hamdi H, Welsh J, Levicar N, Marley SB, Nicholls JP, et al. High frequency of fetal cells within a primitive stem cell population in maternal blood. Hum Reprod 2008; 23(4): 928–33
50. O'Donoghue K, Sultan HA, Al-Allaf FA, Anderson JR, Wyatt-Ashmead J, Fisk NM. Microchimeric fetal cells cluster at sites of tissue injury in lung decades after pregnancy. Reprod Biomed Online 2008;16: 382–90
51. Rossi G. Nature of stem cells involved in fetomaternal microchimerism. Lancet 2004; 364: 1936
52. Khosrotehrani K, Johnson KL, Cha DH, Salomon RN, Bianchi DW. Transfer of fetal cells with multilineage potential to maternal tissue. JAMA 2004; 292: 75–80
53. Srivatsa B, Srivatsa S, Johnson KL, Samura O, Lee SL, Bianchi DW. Microchimerism of presumed fetal origin in thyroid specimens from women: a case-control study. Lancet 2001; 358: 2034–8
54. Oddy WH, Sly PD, de Klerk NH, Landau LI, Kendall GE, Holt PG, et al. Breast feeding and respiratory morbidity in infancy: a birth cohort study. Arch Dis Child 2003; 88: 224–8
55. Chantry CJ, Howard CR, Auinger P. Full breastfeeding duration and associated decrease in respiratory tract infection in US children. Pediatrics 2006; 117: 425–32
56. Duijts L, Jaddoe VWV, Hofman A, Moll HA. Prolonged and exclusive breastfeeding reduces the risk of infectious diseases in infancy. Pediatrics 2010; 126: 1 e18–e25
57. Chisti MJ, Salam MA, Smith JH, Ahmed T, Ashraf H, Bardhan PK, et al. Impact of lack of breast feeding during neonatal age on the development of clinical signs of pneumonia and hypoxemia in young infants with diarrhea. PLoS One 2011; 6: e25817
58. Hoddinott P, Tappin D, Wright C. Breast feeding. BMJ. 2008; 336(7649): 881–7
59. Section on breastfeeding. Breastfeeding and the use of human milk. Pediatrics. 2012; 129(3): e827–41